Research Article
Characterization of Sucrose Synthase (Susy) Transcription Factor from Saccharum officinarum Co86032 during Abiotic Stress Condition
Sreeja PK and Geethalakshmi S*
Department of Biotechnology, Nehru Arts and Science College, India
Corresponding author: Geethalakshmi S, Department of Biotechnology, Nehru Arts and Science College, Coimbatore, Tamil Nadu, India, E-mail: s.geethalakshmi@gmail.com
Citation: Sreeja PK, Geethalakshmi S. Characterization of Sucrose Synthase (Susy) Transcription Factor from Saccharum officinarum Co86032 during Abiotic Stress Condition. J Plant Sci Res. 2019;6(1): 182.
Copyright: © Sreeja PK, et al. 2019. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Plant Science & Research | ISSN: 2349-2805 | Volume: 6, Issue: 1
Submission: 04/02/2019; Accepted: 28/02/2019; Published: 03/05/2019
Abstract
Sugarcane is one of the oldest energy source and currently the major biofuel crop in India. Worldwide, sugarcane is the important source of sugar and the byproducts has several applications. Sucrose gets accumulated in the leaves of sugarcane and is distributed in the culm by various gene mechanisms. The variety of sugarcane Co86032 is the major industrial crop which accumulates high quantity of sucrose. The process of sucrose synthesis is mediated by a family of Transcription Factors (TF) called Sucrose Synthase (SuSy) genes which belong to the MYB transcription factor family. The MYB TF is one of the largest gene family in plants which is involved in almost all the growth and development process of the plants. This family has a special property of getting expressed in the stress condition helping the plant to retain its normal biochemical process, and so it act as stress responsive gene family. This study aims at isolating SuSy gene(s) from Saccharum officinarum Co86032 during abiotic stress condition like drought and salt stress and characterizing its expression level using RT-PCR. The expression level of the amplified gene in all the stress samples was analyzed and it was found that the DNA got expressed at almost the same level even at the 20th DOS (Day of Stress). Thus, this study forms a basis for identification of the sucrose synthase gene which has high level of expression during abiotic stress condition.
Keywords:
MYB; Sugarcane; abiotic stress; Sucrose synthase; Transcription factor
Introduction
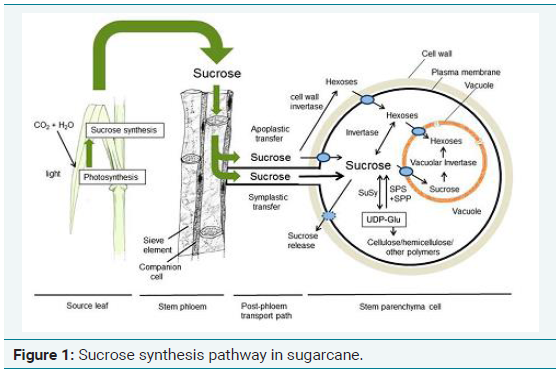
Sugarcane (Saccharum officinarum L.), is an old energy source for human beings and more recently, a replacement of fossil fuel for motor vehicles. The current cultivars of sugarcane are highly polyploid and aneuploid and contains on an average of 100-120 chromosomes with cell size of 10,000 Mbp [1]. Significant progress has been noted recently with the development of Expressed Sequence Tags (EST). Generally, plants face biotic stresses like pests, pathogens, rodents and abiotic stresses like drought, flood, salinity, extreme temperature, high light, deficit in nutrients and heavy metal toxicity [2]. Among the many stress factors, abiotic stress can directly or indirectly affect the physiological status of the plant by altering its metabolism, growth and development. The sucrose synthesis pathway in sugarcane is shown in Figure 1 [3]. Sucrose Synthase (SuSy) is one of the key enzymes involved in sucrose metabolism, catalyzing the reversible conversion of sucrose and UDP to UDP-glucose and fructose. Therefore, its activity, localization and function have been studied in various plant species. It has been shown that SuSy can play a role in supplying energy in companion cells for phloem loading [4], provides substrates for starch synthesis [5], and supplies UDP-glucose for cell wall synthesis nitrogen fixation and response to abiotic stresses [6]. In order to increase the sucrose content at the molecular level, it is very necessary to understand the biological function of sucrose synthase gene in sugarcane. In sugarcane, MYB TF family is majorly involved in sucrose synthesis even under stress conditions [7,8].
With these importances of sugarcane and SuSy TFs, the present study aims at identifying the SuSy gene homologues in sugarcane using the available EST data and by comparing the EST sequences with other plant genes. Primers specific to these genes are designed using computational approaches. Further, in order to analyze the expression of these predicted genes in stress conditions; qRT-PCR analysis was done for the SuSy genes isolated from leaf tissues of sugarcane.
Materials and Methods
Plant material
Setts of sugarcane (Saccharum sp.) plants of cultivar, Co86032 in the form of entire transverse nodal culm sections, bearing a single intact axillary bud was collected from Sugarcane Breeding Institute, Coimbatore, Tamil Nadu, India. The collected plant materials were maintained in plastic pots containing soil: vermiculite: organic fertilizer, 3:2:1, (w/w/w) under greenhouse conditions (16/8-h photoperiod; 26 °C). The sugarcane plantlets developed from the axillary bud of the setts were used for stress induction.
To identify sucrose synthase (SuSy) gene homologues in sugarcane by computational methods, the steps indicated in were followed and specific primers for the SuSy genes were designed using Primer-BLAST (http://www.ncbi.nlm.nih.gov/tools/primer-blast/primerinfo.html) [7].
Stress induction in sugarcane plantlets
The setts of sugarcane maintained in greenhouse conditions germinated after 8 days of planting. The plantlets were divided into three groups of three plants each; one group served as control, the second group was induced with drought stress and the third group was induced with salt stress. To develop water-deficit stress, watering to the test plants were withheld during the tillering stage (30 - 60 days after planting) for the designated period. For salt stress, test plants were grown in soil containing 2 % (w/w) of NaCl. The control plants were maintained without salt stress and received normal watering throughout the experiment. The uppermost fully expanded leaf samples were collected at 0, 4, 8, 12, 16 and 20 Days after Stress (DOS) from control, water-deficit and salt stressed plants. The collected leaf tissues were quick-frozen and stored at -80 °C until RNA extraction. RNA was extracted from the leaf samples using RNA extraction and purification kit (Sigma Aldrich) as per manufacturer’s instructions and cDNA was synthesized.
Amplification of the SuSy gene coding sequence
The cDNA synthesized from the leaf samples were subjected to PCR amplification. A total of 9 primers were designed for SuSy gene using Primer-BLAST were used for amplification of the synthesized DNA (Both Control and stress induced) using PCR (Millipore) as per manufacturer’s instructions. The unincorporated PCR primers and dNTPs were removed from PCR products by using Montage PCR Clean up kit (Millipore). The PCR product was sequenced using the primers. Sequencing reactions were performed using ABI PRISM® BigDyeTM Terminator Cycle Sequencing Kits with AmpliTaq® DNA polymerase (FS enzyme) as per manufacturer’s instructions (Applied Biosystems). The sequenced genes were subjected to Bioinformatics analysis.
Bioinformatics Protocol
1. The 16s rRNA sequence was blast using NCBI blast similarity search tool. The phylogeny analysis of query sequence with the closely related sequence of blast results was performed followed by multiple sequence alignment.
2. The program MUSCLE 3.7 was used for multiple alignments of sequences [9]. The resulting aligned sequences were cured using the program Gblocks 0.91b. This Gblocks eliminates poorly aligned positions and divergent regions (removes alignment noise) [10]. Finally, the program PhyML 3.0 aLRT was used for phylogeny analysis and HKY85 as Substitution model.
3. PhyML was shown to be at least as accurate as other existing phylogeny programs using simulated data, while being one order of magnitude faster. PhyML was shown to be at least as accurate as other existing phylogeny programs using simulated data, while being one order of magnitude faster. The program TreeDyn 198.3 was used for tree rendering [11].
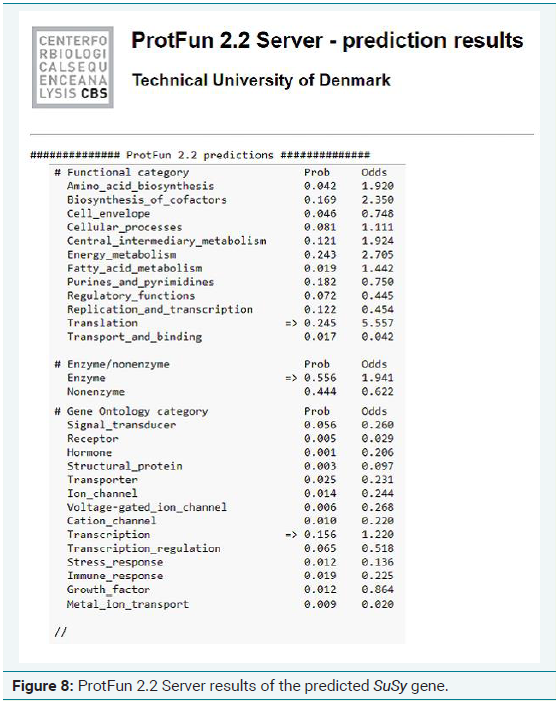
4. The protein sequences of the identified genes were further subjected to ProtFun 2.2 server (http://www.cbs.dtu.dk/services/ProtFun/) to predict the function of the identified sequences. The sequences of the identified motifs were given as the input in the FASTA format for the predicting their functions. The parameters were set as default.
Expression analysis of the identified SuSy gene using RTPCR during abiotic stress condition
To validate the predicted function and expression pattern of the predicted SuSy genes, RT-PCR was performed by following the established procedures using the isolated RNA samples and designed primers [12]. Real Time - PCR (RT-PCR) amplification was performed using a MX3005P thermocycler (Stratagene/ Agilent Technologies) as per standard protocol. The RT reaction mix without reverse transcriptase served as a negative control. For internal control, primers NtActin_FP: 5’-ATGGCAGACGGTGAGGATATTCA-3’ and NtActin_RP: 5’-GCCTTTGCAATCCACAT CTGTTG-3’ was used for amplification.
The dissociation curves were generated at 95 °C to verify the PCR specificity after completion of PCR cycling. Quantities of RNA accumulation levels were calculated as RQ values using the comparative cycle threshold (CT) (2-ΔΔCt) method. Dissociation curve/ Melt curve is generated by increasing the temperature from 55 °C to 95 °C in tiny increments and monitoring the intensity of fluorescence at each step. All qPCR reactions were performed in three technical replicates.
Results and Discussion
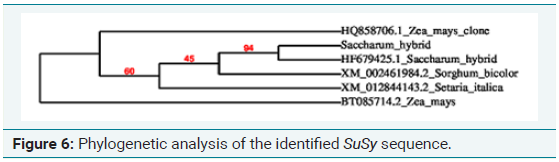
Comparative genomics studies have been used to analyze SuSy gene families in various plant species, including citrus, tobacco, poplar, and cotton [13,14]. Since the complete genome of sugarcane is not available as it is polyploidy in nature and is very difficult to determine the complete CDS regions, the only option left behind is the usage of EST sequences. According to the computational analysis, SuSy gene homologues were searched in sugarcane nucleotide sequences. SuSy genes (896 genes) collected from plant transcription databases were subjected to local BLAST against the 3268 sugarcane transcription factor genes involved in sucrose synthesis. A total of 75 putative sequences were identified after the local BLAST analysis. After redundancy removal, the individual sequences were subjected to contig assembly using the CAP3 program which resulted in 26 candidate sequences. The resulting sequences were subjected to PROSITE, MEME and ProtFun analysis program to confirm their characteristic domain and its function [15].
Abiotic stress causes many changes in the morphological, physical and biochemical properties of a plant which may lead to decrease in the regular metabolic activities of the plant (Figure 2). The morphological changes start with decrease in the chlorophyll content, curling of the tips of the leaf, dryness, yellowness of the leaf and finally to turning the leaf to brown color loosing the rigidity. These changes occur slowly at a constant rate under salt stress as well as drought stress conditions [16]. The additional changes that can be observed in drought stress is change in the osmotic conditions of the plant leading to decrease in the relative water content of the plant. This change affects the photosynthesis of the plant which is connected with the sucrose synthesis and transport [3], since sucrose in synthesized during the photosystem II [17], gets transported to the culm of the sugarcane and accumulates there. The quantity of accumulated sucrose decides the length and thickness of the stalk of the sugarcane which decides its commercial value.

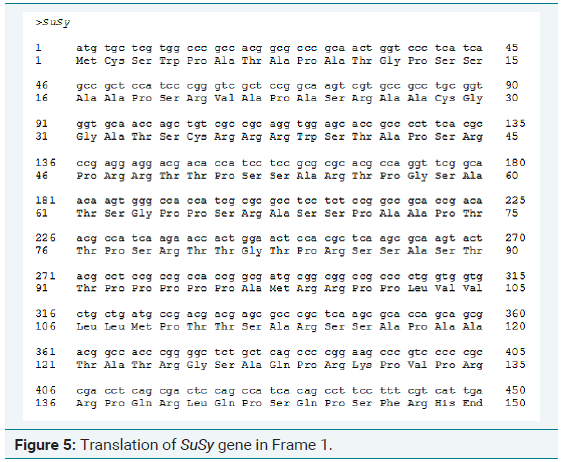
Of the 9 primers designed, only one gave a good result which is shown in Table 1. So, this primer was taken for further expression analysis in the stress induced samples. The amplified PCR product was quantified, purified and sequenced to obtain the complete sequence of the amplified gene (Figure 3). From the Figure 3, it is clear that the primer designed for analyzing the synthesis of sucrose synthase during abiotic stress conditions gave a good amplification the possibility of expression of the SuSy gene during stress condition also. The nucleotide sequence thus obtained was subjected to Bioinformatics analysis. Figure 4-8 depicts the results of Bioinformatics analysis which confirms the isolated gene is SuSy gene belonging to MYB transcription factor family.
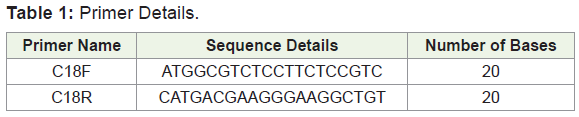

Figure 3: PCR amplification of DNA from stress induced samples using designed primer.
C - Control; D4 - Day 4, D8 - Day 8, D12 - Day 12, D16 - Day 16, D20 - Day 20 of stress induction. Drought - DNA isolated from drought stress induced leaf samples; Salt - DNA isolated from salt stress induced leaf samples.


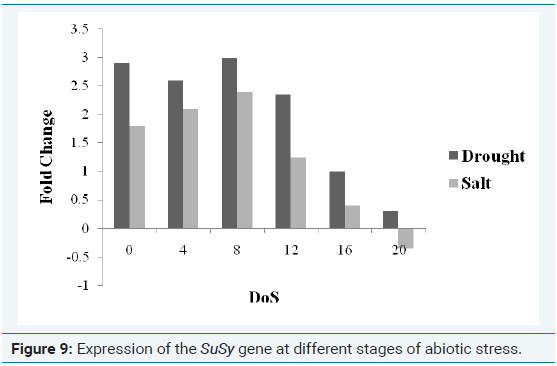
Expression patterns of stress inducible genes are very complex. Some genes respond to drought, salt and cold very rapidly, whereas others are induced slowly after the accumulation of hormonal signals. RT-PCR was carried out using primers specific to SuSy gene for validating its predicted biological function. In this study, Sc25S rRNA was used as reference gene for RT-qPCR experiment [18]. The SuSy transcripts expression level at different stages of stress is indicated in Figure 9. From the above figure, it can be understood that the SuSy gene got expressed at almost same level during the initial period of stress (0-8 DOS) and from 12th DOS, the level of expression started decreasing. Interestingly, on the 20th DOS, it is found that the SuSy gene got under expressed in the drought condition. This can be correlated with the non availability of water on the 20th DOS which seriously affects the photosynthesis of the plant. Since the light reaction of photosynthesis cannot occur in the absence of water, the synthesis process of sucrose drastically decreased as the gene was not able to express itself.
Summary
The results of the study can be summarized as follows: 3268 sucrose synthase genes from sugarcane collected from transcription factor databases were analyzed computationally and 26 unique candidate genes were shortlisted. These 26 genes were overlapped for complete CDS formation and 9 gene specific primers were designed. Saccharum officinarum Co86032 was maintained in the green house condition and abiotic stress (Salt stress and Drought) was induced; leaf samples were collected at 4 days interval, RNA was isolated, cDNA synthesized and amplified using gene specific primers. Only one primer gave good amplification in all the samples and so that amplified gene was taken for sequencing. The sequenced gene and identified protein sequence was subjected to various Bioinformatics analysis to identify the motifs and its functional role. The gene was taken for RT-PCR analysis and the level of expression of the gene wa analyzed in the stressed as well as control plants. The identified SuSy gene was expressed at a reasonable level during the initial period of stress but the expression level started decreasing after 12th DOS. It can be inferred that this gene which got expressed during the initial period has the capacity to withstand the abiotic stress to certain level. If this candidate gene is taken for overexpression studies, it can serve as a potential candidate for developing transgenic sugarcane which contains the overexpressed SuSy gene incorporated into its genome which will render resistance for the plant against abiotic stress. Also, during the stress condition, if the expression level of a gene remains standard, the gene will be able to induce the synthesis of sucrose. Thus, this study will form a basis for gene identification and expression and developing stress resistant sugarcane plant which accumulates high sucrose content even under stress conditions and development of transgenic plants containing overexpressed gene will be beneficial for industrial production of sugar.
Acknowledgements
The authors are thankful to UGC-SERO for funding this work.
References
- D’Hont A, Glaszmann JC (2001) Sugarcane genome analysis with molecular markers, a first decade of research. Proc Int Soc Sugarcane Technol 24; 556-559.
- Yu Y, Song C, Zhang Q, DiMaggio PA, Garcia BA, et al. (2012) Histone H3 lysine 56 methylation regulates DNA replication through its interaction with PCNA. Mol Cell 46: 7-17.
- Zhu YJ, Komor E, Moore PH (1997) Sucrose accumulation in the sugarcane stem is regulated by the difference between the activities of soluble acid invertase and sucrose phosphate synthase. Plant Physiol 115: 609-616.
- Fu H, Park WD (1995) Sink- and vascular-associated sucrose synthase functions are encoded by different gene classes in potato. Plant Cell 7: 1369-1385.
- Zrenner R, Salanoubat M, Willmitzer L, Sonnewald U (1995) Evidence of the crucial role of sucrose synthase for sink strength using transgenic potato plants (Solanum tuberosum L.). Plant J 7: 97-107.
- Haigler CH, Ivanova-Datcheva M, Hogan PS, Salnikov VV, Hwang S, et al. (2001) Carbon partitioning to cellulose synthesis. Plant Mol Biol 47: 29-51.
- Geethalakshmi S, Barathkumar S, Prabu G (2015) The MYB transcription factor family genes in sugarcane (Saccharum sp.). Plant Mol Biol Rep 33: 512-531.
- Iskandar HM, Casu RE, Fletcher AT, Schmidt S, Xu J, et al. (2011) Identification of drought-response genes and a study of their expression during sucrose accumulation and water deficit in sugarcane culms. BMC Plant Biol 11: 12.
- Edgar RC (2004) MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res 32: 1792-1797.
- Talavera G, Castresana J (2007) Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst Biol 56: 564-577.
- Dereeper A, Guignon V, Blanc G, Audic S, Buffet S, et al. (2008) Phylogeny. fr: robust phylogenetic analysis for the non-specialist. Nucleic Acids Res 36: W465-W469.
- Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25: 402-408.
- Islam MZ, Hu XM, Jin LF, Liu YZ, Peng SA (2014) Genome-wide identification and expression profile analysis of citrus sucrose synthase genes: investigation of possible roles in the regulation of sugar accumulation. PLoS One 9: e113623.
- Wang W, Vinocur B, Shoseyov O, Altman A (2015) Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci 9: 244-252.
- Erpen L, Devi HS, Grosser JW, Dutt M (2018) Potential use of the DREB/ERF, MYB, NAC and WRKY transcription factors to improve abiotic and biotic stress in transgenic plants. Plant Cell Tiss Organ Cult 132: 1-25.
- Zlatev Z, Lidon FC (2012) An overview on drought induced changes in plant growth, water relations and photosynthesis. Emir J Food Agric 24: 57-72.
- Suresha GS, Mahadevaiah C, Appunu C (2017) Biotechnological interventions for improving sucrose accumulation in sugarcane. In: Mohan C (eds.) Sugarcane Biotechnology: Challenges and Prospects. Springer, Cham pp: 111-122.
- Gibson UE, Heid CA, Williams PM (1996) A novel method for real time quantitative RT- PCR. Genome Res 6: 995-1001.