Research article
Comparative Wood Anatomy of 37 Woody Species of Tamaulipan Thorn Scrub, Northwestern Mexico and Its Relation to Adaptation and Wood Quality: A Synthesis
Maiti R and Rodriguez HG*
Universidad Autónoma de Nuevo León, Facultad de Ciencias Forestales, México
Corresponding author: Rodriguez HG, Universidad Autónoma de Nuevo León, Facultad de Ciencias Forestales, Carr. Nac. No. 85 Km. 45, Linares, Nuevo León 67700, México, E-mail: humberto.gonzlez@uanl.mx
Citation: Maiti R, Rodriguez HG. Comparative Wood Anatomy of 37 Woody Species of Tamaulipan Thorn Scrub, Northwestern Mexico and Its Relation to Adaptation and Wood Quality: A Synthesis. J Plant Sci Res. 2018;5(1): 175.
Copyright © Maiti R 2018. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Plant Science & Research | ISSN: 2349-2805 | Volume: 5, Issue: 1
Submission: 21/12/2017; Accepted: 25/01/2018; Published: 31/01/2018
Abstract
The paper gives a brief account of wood anatomy of 37 woody species of Tamaulipan Thorn Scrub at Linares, on the basis of transverse, tangential sections and maceration of fiber cells and xylem vessels characteristics. Some of these results are published. In transverse sections there exists large varia variations in distribution of xylem vessels, varying in size, arrangement in radial bands, groups or isolated which are the characteristics of the particular species, inter-specific. Many of the species possess narrow vessels which are reported by authors against occlusion owing to drought or cold, an adaptive trait. Some species poses high density of thick walled lignified fiber cells which I related to the production of strong furnitures, few having soft fibre cells which may be related to paper quality. In transverse sections, the species show variation in the type of axial parenchyma, paratracheal, apotracheal, etc, characteristics of the species. In tangential sections the species show variation in the types of ray parenchyma, uniseriate, long or short or multiseriate , long and short, Fiber cells show variation among species in the length, breadth, cell wall thickness which may be related to taxonomic determination and quality determinations of the species. Vessels show variation in length, breadth, inclination of end walls which may be related to evolutionary trends and taxonomic determination of the species. Therefore, wood anatomy plays an important role in taxonomic delimitation of the species, adaptation to environmental stresses and quality of wood and its utility.
Keywords:
Wood anatomy; Tamaulian woody species; Variations; Vessel size; Shape; Distribution; Types of axial parenchyma; Wood p rays; Fiber cell length; Breadth; Wall thickness; Taxonomic determination; Wood quality
Introduction
Wood is an important product of woody plants for wood industry and used for furniture and building construction also used for fire wood. Significant research advances have been undertaken on wood anatomy and its significance in dendrology and application. Few studies have been undertaken on ultrastructural and biochemical changes in the development of wood elements.
A study on the vessel and fibre orientations in Acacia Mangium Willd. Both vessel and fiber orientations showed similarity in radial characteristics and distinct inversion of the grain. But, the vessel orientation showed larger amplitude of change than fiber orientation [1].
The secondary wall structure of tension wood of Laetia procera Poepp. (Flourtiaceae), revealed alternate arrangement of thick and thin layers with S1 + S2 + S3 using UV microspectrophotometry. It was observed that in the thick secondary wall, cellulose microfibril angle is very low (very close to fibre axis) and cellulose microfibrils are well organized but in thin layer the cellulose microfibrils are less organized and oriented with a large angle in the axis of the cell. Thick layers are highly lignified [2].
Later, a study undertaken on wood anatomy and ultra structure of the 3 species of wood of Prosopis, Prosopis vinalillo, Prosopis alba and Prosopis nigra in heterogeneous forest dry Chaqueno Park revealed that the 3 species showed similarity in the structural features of the subfamily Mimosoideae. However, the number of vessels/mm2 showed large variations among species and between individuals of the same species. Using scanning electron microscope the ornaments in pits and striations on the vessels walls were observed. The variation in striations was observed to be characteristics of in the 3 Prosopis species [3].
A study on wood anatomy of Prosopis pallida in the arid and semi-arid lands of the American continent revealed that P. pallida has well-differentiated annual growth rings which are related well with precipitation events related to El Nino Southern Oscillation phases [4].
Wood anatomy is used to determine the specific characteristics of species.
A comparative study undertaken on macroscopic and microscopic anatomical characteristics of five species of the family Rosaceae, Crataegus mexicana, Pyrus communis, Pyrus malus, Prunus americana and Prunus domestica, showed similar macro and microscopic characteristics [5]. There exists a large variability in size, cell wall thickness and lumen breadth which may predict the quality and utility of the particular species [6].
Wood anatomical traits are found to be related to the adaptation of woody plants to environmental stresses.
A study was undertaken on the anatomical heartwood. It was observed that in the locality Linares, Nuevo Leon, Mexico, with higher precipitation and lower temperature the wood showed higher fibre length and higher diameter of the vessels than China, Nuevo Leon [7].
In addition to the basic studies on the growth and development of wood elements, wood anatomical features play important role in the phylogeny of the species and also the adaptive capacity of the species to environmental stresses [8-12]. The hydraulic architecture of woody plants determines the adaptive strategies to adverse climatic conditions of woody plants [13-19]. From a functional viewpoint, few vessel attributes such as narrow pores and pores multiples acts against cavitation and embolism under hot summer and freezing stress, thereby offering mechanical strength [14,20-23].
Few studies have been undertaken on wood anatomy of Mediterranean woody species in relation to ecology and ecophysiology.
Various authors stated that the presence of narrow vessels and multiple vessels acts against cavitation during summer stress and winter freezing. Sperry, studied patterns in hydraulic architecture and their implications for transport efficiency [24].
The stem and root wood anatomy of the shrub-Phlomis Fruticosa (Labiatae) a malacophilous Mediterranean drought semi-deciduous species has shown that the stem is comprised of diffuse-porous [25], narrow vessels arranged in tangential bands, vessel elements with oblique simple perforation plates, non-vestured, clustered alternate intervessel pits. It is concluded that though narrow vessels offer high conducting resistance, they are less vulnerable to cavitations, thus providing safety during summer drought and winter freezing. Vessel grouping is a widespread phenomenon in most woody species, especially those from the arid desert flora and Mediterranean species [26].
Veronica De Micco, et al. studied wood anatomy and hydraulic architecture of stems and twigs of some Mediterranean trees and shrubs along a mesic-xeric gradient [27]. This study focuses on the anatomy of juvenile and mature wood of some species representative of continuous sequences of Mediterranean vegetation formations according to gradients of water availability, from xeric to relatively mesic: Although some attributes (i.e. porosity and type of imperforate tracheary elements) were similar in young twigs and older rings, other traits (i.e. vessel frequency and size) revealed the different hydraulic properties of twig and stem wood. The difference between juvenile and mature structures was large in the species of the mesic end of the gradient while it was relatively small in those more xeric. The species showed large variations in wood anatomical traits, most of them are diffuse porous, few semi to ring porous, vessels are narrow resistant to cavitation during drought and freezing.
In Mediterranean-type ecosystems, seasonal dimorphism is an adaptive strategy to save water by developing brachyblasts with xeromorphic summer leaves as opposed to dolicho blasts with more mesomorphic winter leaves. A study was undertaken on the seasonal dimorphism in wood anatomy studied in Mediterranean sub sp Cistus incanus has shown that brachyblast wood was safer than dolicho blast and has narrower and more frequent vessels [27]. The measurement of other specific anatomical traits, such as vessel wall thickness, suggested that brachyblast wood has a higher resistance to implosion due to drought-induced embolism.
Methodology
Transverse and tangential and radial sections of woods of 3t woody species have been undertaken according to standard protocols and maceration of woods have been undertaken according to Maiti, et al. for fibre cells and vessels [6].
Results and Discussion
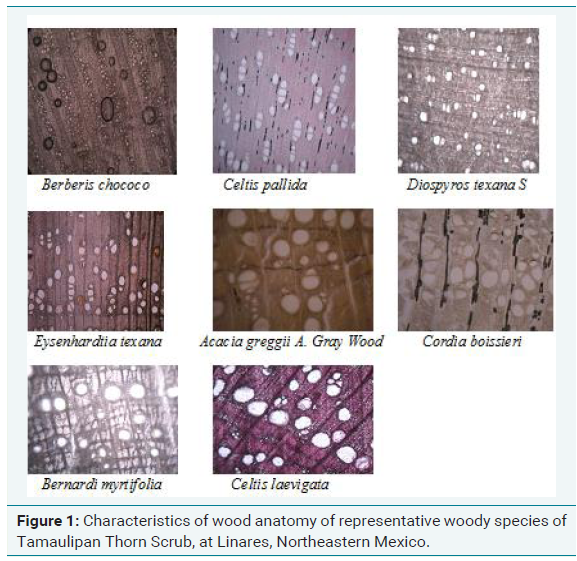
A study undertaken by Maiti, et al. on comparative anatomy of 20 woody species of Tamaulipan Thorn Scrub revealed that a study has been undertaken on wood anatomy of 20 woody species in northeast Mexico [28]. There exists large variation among species in wood anatomical traits such as porosity, vessel diameter, its distribution, parenchyma, compactness of ground tissues and fibre cell characteristics. Most of the species have narrow vessels which help protection against cavitation. Most of the species are ring to semi ring porous viz. Acacia amentacea, Acacia berlandieri, Acacia schaffneri, Acacia wrightii, Cordia boissieri, Helietta parvifolia, Condalia hookeri, Xanthoxylum fagara, Celtis pallida, Celtis laevigata, Caesalpinia mexicana, Eysenhardtia polystachya; only few of them are diffuse porous viz. Diospyros palmeri, Diospyros texana. Fibre cell characteristics also showed large variations in morphology, size, lumen breadth and in compactness, these species were soft and hard wood. Most of the species have narrow vessels, viz., Acacia berlandieri, Acacia schaffneri, Acacia wrightii, Helietta parvifolia, Cordia boissieri, Diospyros palmeri, Celtis laevigata, Eysenhardtia polystachya, Zanthoxylum fagara. Celtis pallida contained medium sized vessels, while Celtis laevigata and Caesalpinia mexicana possessed big sized vessels. Many of the species possess narrow vessels which although impose transport of water but protect the vessels against cavitation during drought and freezing the species with small narrow vessels mentioned have strategy to adapt both to hot and cold climate against cavitation. The species having big vessel diameter may be susceptible to drought such as Celtis pallida, Caesalpinia mexicana or they may have deep root system for adaptation to semiarid climates in northeast Mexico. All these wood anatomical traits could be utilized to distinguish species as well as quality determinations of species. The variation in hydraulic systems determines the capacity of water transport among species statistically significant differences are observed in all wood anatomical parameters among species studied.
In the present study, we undertook characterization of wood anatomy of 37 woody species of Tamaulipan Thorn Scrub. Most of the species possess diffuse porous woods and very few ring porous. Many species possess narrow vessels. We observed large variation among 37 woody species in wood anatomical traits among woody species with respect type, size of vessels (long, narrow, short, truncated, inclined etc.), wood porosity, vessel density, type of wood parenchyma (paratracheal, vescicentric, apotracheal, alliform etc), vessels may be isolated, solitary, in radial bands or ingroups, ray cells (uniseriate, multiseriate, short or long), vessel pitting, wood fibre cell characteristics, wood compactness which could be utilized in taxonomic delimitation, wood quality and utility. A study was undertaken by Maiti, et al. to determine the variability in fiber cell morphology and its length among wood species of the Tamaulipan thornscrub, northeastern Mexico, used for various uses such as timber for furniture, fence, post, firewood and sources of forage [6]. The results reveal large variability in fiber cell morphology and fiber cell length. The species have been classified on the basis of its fiber cell morphology and fiber cell length and recommended for their possible utilization for different purposes. Species with highly lignified fiber cell walls are suitable for strong furniture while those with thin wall for paper making.
In the semiarid hot summer many species possess narrow vessels and high xylem density which could function against cavitation and embolism against hot summer and cold seasons which have been reported by various authors (the adaptive capacity of the species to environmental stresses [8,10,11]. Species with high xylem density are reported to be drought tolerant [23].
We selected species with high xylem density and narrow vessels. Acacia wrightii, Berberis chococo, Diospyros pallens, Diospyros texana, Ehretia anacua, Fraxinus greggii, Guia cum angustifolia, Helietta parvifolia, Karwinskia humboldtiana and others. Research needs to be directed to verify the capacity of these species against cavitation and drought tolerance (Figure 1).
Conclusion
This paper presents a short summary of comparative characterization of 37 species in Northeastern Mexico revealing large variability in wood anatomical structures. The species have been grouped in distinct categories on the basis of specific characteristics. It is suggested that these variations could be used in taxonomic delimitation of the species as well as in the adaptation of the species in xeric habitats in summer and cool environments in winter. Research inputs should to be directed to relate wood anatomical traits with their adaptations to hot and cold climates.
References
- Ogata Y, Fujita M, Nobuchi T, Shri MH (2003) Macroscopic and anatomical investigation of interlocked grain in Acacia mangium. IAWA Journal 24: 13-26.
- Ruelle J, Yoshida M, Clair B, Thibaut B (2007) Peculiar tension wood structure in Laetia procera (Poepp.) Eichl. (Flacourtiaceae). Trees 21: 345-355.
- Muniz BDGI, Nisgoski S, Lonelí-Ramírez G (2010) Wood anatomy and ultrastructure of three species of Prosopis (Leguminosae-Mimosoideae) from dry Parque Chaqueno, Argentina. Madera y Bosquees 16: 21-38.
- López BC, Sabaté S, Gracia CA, Rodríguez R (2005) Wood anatomy, description of annual rings, and responses to ENSO events of Prosopis pallida H.B.K., a wide-spread woody plant of arid and semi-arid lands of Latin America. J. Arid Environ 61: 541-554.
- Olvera CP, Aguirre MM, Romero JC, Pacheco YL (2008) Wood anatomy of five specie of the Rosaceae family. Madera y Bosque 14: 81-105.
- Maiti R, Para AC, Rodriguez HG, Paloma SV (2015) Characterization of wood fibres of scrubs and tree species of the tamaulipan thornscrub, northeastern mexico and its possible utilization. Forest Res 4: 1-5.
- Parra AC, Pournavab RF, García VB, Torres SS, Ocanas FG, et al. (2013) Differences of wood elements of Prosopis laevigata from two areas of Northeast Mexico. Am J Plant Sci 4: 56-60.
- Bailey IW, Tupper WW (1918) Size variation in tracheary cells. I. A comparison between the secondary xylem of vascular cryptogams, gymnosperms and angiosperms. Proc Am Acad Arts Sci 54: 149-204.
- Carlquist S (1984) Vessel grouping in dicotyledon wood: significance and relationship to imperforate tracheary elements. ALISO 10: 505-525.
- Baas P (1982) Systematic, phylogenetic and ecological wood anatomy- History and perspectives. In: Baas P (edn.) New Perspectives in Wood Anatomy. Nijhoff/Junk, The Hague pp. 23-58.
- Baas P, Ewers FW, Davis SD, Wheeler EA (2004) Evolution of xylem physiology. In: Hemsley A, Poole I (edn.) The evolution of plant physiology. Elsevier pp. 273-295.
- Ewers FW, JB Fisher (1991) Why vines have narrow stems: Histological trends in Bauhinia (Fabaceae). Oecologia 88: 233-237.
- Carlquist S (1989) Adaptive wood anatomy of chaparral shrubs. In: Keely JE (edn.) The California chaparral: paradigms re-examined. Los Angeles Country. Museum of Natural History Contributions, Los Angeles pp. 25-35.
- Zimmermann MH (1982) Functional xylem anatomy of angiosperm trees. In: Baas P (edn.) New perspectives in wood anatomy. Xylem Structure and the Ascent of Sap. Springer, Berlin pp. 59-70.
- Baas P, Carlquist S (1985) A comparison of the ecological wood anatomy of the floras of Southern California and Israel. IAWA J 6: 349-353.
- Baas P, Schweingruber FH (1987) Ecological trends in the wood anatomy of trees, shrubs and climbers from Europe. IAWA J 8: 245-274.
- Tyree MT, Ewers FW (1991) The hydraulic architecture of trees and other woody plants. Transley review no. 34. New Phytol 119: 345-360.
- Tyree M, Davis SD, Cochard H (1994) Biophysical perspectives of xylem evolution: is there a tradeoff of hydraulic efficiency for vulnerability to dysfunction? IAWA J 15: 335-360.
- Hacke UG, Sperry JS, Pockman WT, Davis SD, MCculloh KA (2001) Trends in wood density and structure are linked to prevention of xylem implosion by negative pressure. Oecologia 126: 457-461.
- Ewers FW (1985) Xylem structure and water conduction in conifer trees, dicot trees, and lianas. IAWA J 6: 309-317.
- Salleo S, MA Lo Gullo (1994) Drought resistance strategies and vulnerability to cavitation of some Mediterranean sclerophyllous trees. In: Borghetti M, Grace J, Rasch A (edn.) Water transport in plants under climatic stress. Cambridge University Press, Cambridge pp. 99-113.
- Hacke UG, Sperry JS, Wheeler JK, Castro L (2006) Scaling of angiosperm xylem structure with safety and efficiency. Tree Physiol 26: 689-701.
- Jacobsen AL, Agenbag L, Esler KJ, Pratt RB, Ewers FW, et al. (2007) Xylem density, biomechanics, and anatomical traits correlate with water stress in 17 evergreen shrub species of the Mediterranean-type climate region of South Africa. J Ecology 95: 171-183.
- Psaras GK, Sofroniou I (2004) Stem and root wood anatomy of the shrub-Phlomis fruticosa (Labiatae). IAWA J 25: 71-77.
- McCulloh KA, Sperry JS (2005) Patterns in hydraulic architecture and their implications for transport efficiency. Tree Physiol 25: 257-267.
- Fahn A, Er EW, Baas P (1986) Wood anatomy and identification of trees and shrubs from Israel and adjacent regions. In: Miller RB The Israel Academy of Sciences and Humanities, Jerusalem. IAWA J 7: 221.
- Micco VD, Aronne G, Bass P (2009) Wood anatomy and hydraulic architecture of stems and twigs of some Mediterranean trees and shrubs along a mesic-xeric gradient. Trees 22: 643-655.
- Ratikanta M, Rodriguez HG, Jeff Christofher GD, Alejandra DT, Allejandro CF, et al. (2016) Comparative wood anatomy of 20 woody plant species in Northeastern Mexico and its significance. Int J of Bioresource and Stress Manag 7: 229-246.