Research Article
Interspecific Difference in Boron tolerance at Seed Germination Stage in three Hibiscus species
Babu A Sonar1*, Dattatray K Gaikwad2, Sunil D Kamble3 and Prakash D Chavan4
1,3Department of Botany, Arts, Science and Commerce College, Ramanandnagar (Burli) (MS), India
2,4Department of Botany, Shivaji University, Kolhapur (MS), India
Corresponding author: Babu A Sonar, Department of Botany, Arts, Science and Commerce College,Ramanandnagar (Burli) (MS), India-416 004, Tel: +91 9860121992; E-mail: baburao.2007sonar@gmail.com
Citation: Sonar BA, Gaikwad DK, Kamble SD, Chavan PD. Interspecific Difference in Boron tolerance at Seed Germination Stage in three Hibiscus species. J Plant Sci Res. 2016;3(2): 154.
Copyright © Sonar et al., 2016. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Plant Science & Research | ISSN: 2349-2805 | Volume: 3, Issue: 2
Submission: 23/06/2016; Accepted: 18/07/2016; Published: 23/07/2016
Abstract
Influence of boron toxicity on seed germination, seedling growth and activities of two antioxidant enzymes in three Hibiscus species namely, Hibiscus cannabinus L., Hibiscus sabdariffa L. and Hibiscus tiliaceus L. was investigated. Germination percentage of all the three Hibiscus species was decreased due to higher ‘B’ concentration (200 ppm). Higher dose of boron (200 ppm) caused marked reduction in germination percentage in H. tiliaceus (86%) and H. cannabinus (20%) while, in case of H. sabdariffa (2%) decline in germination percentage was evident. Root length of all the three Hibiscus species wasdeclined due to higher concentration of boron (200 ppm). Root length of the three Hibiscus species was affected under ‘B’ stress in the order H. cannabinus> H. sabdariffa > H. tiliaceus. Shoot length was also similarly reduced due to 200 ppm ‘B’ concentration but the order of decrease was H. tiliaceus > H. sabdariffa > H. cannabinus. Higher dose (200 ppm) of boron concentration reduced the total seedling length in all three Hibiscus species. The percent reduction in seedling length at 200 ppm ‘B’ concentration was in the order H. tiliaceus > H. sabdariffa > H. cannabinus. Considerable reduction in vigour index of all the three Hibiscus species was noticed in response to 200 ppm ‘B’ treatment in following order H. tiliaceus > H. cannabinus > H. sabdariffa.Decrease in fresh weight per seedling of H. cannabinus and H. sabdariffa was noticed under ‘B’ toxicity while, in H. tiliaceus there was slight increase in fresh weight at 100 ppm ‘B’ concentration and thereafter it was decreased. Catalase and peroxidase activities in seedlings of H. sabdariffa were greatly enhanced due to 150 ppm and 200 ppm ‘B’ treatment respectively. Lower dose (100 ppm) caused slight increase in catalase activity of both H. cannabinus and H. tiliaceus seedlings. The activity of enzyme peroxidase was reduced in seedlings of H. cannabinus and H. tiliaceus over entire range of ‘B’ treatment. Thus an interspecific difference in boron tolerance at germination stage was evident in the three Hibiscus species.
Keywords: Germination; Hibiscus; Boron toxicity
Introduction
Boron toxicity is an important disorder which can limit plant growth on soils of arid and semi arid environments throughoutworld [1]. High concentrations of B may occur naturally in the soil or in groundwater and such sites have been noticed in Australia, North and South America, Asia, South Africa, Mediterranean and East Europe [1,2]. Boron deficiency can be corrected with the help of certain anthropogenic factors, such as irrigation water containing excess boron, the use of fly ash as an ameliorant in agriculture, boron fertilizers and excessive boron accumulation in soil may caused due to surface mining [1,2]. The occurrence of boron toxicity in Australian soils is recognized as factors limiting cereal (wheat and barley) production [3]. Furthermore, soil salinity is reported to aggravate the negative impact of boron on different agricultural plants [4]. On this back ground screening of various plant species and varieties for boron tolerance is essential to select boron tolerant species. In the present investigation an attempt has been made to study boron tolerance at seed germination stage in three Hibiscus species namely H. cannabinus, H. sabdariffa and H. tiliaceus. Out of these species the H. cannabinus is a fibre crop. H. sabdariffa and H. cannabinus are also leafy vegetables. H. tiliaceus is a halophytic tree species with ornamental value.
Material and Methods
The seeds of local cultivar of H. cannabinus were collected fromlocal farmers while, seeds H. sabdariffa cultivar AS-73-CP-560were obtained from University of Agricultural Sciences, Dharwad(Karnataka). The seeds of H. tiliaceus were collected from estuarineareas in Ratnagiri.
The healthy viable seeds of the three species were sorted out andtreated with 0.1% mercuric chloride for 5 min. for surface sterilization.The seeds were rinsed with distilled water for 4-5 times. Prior to thestart of experiment, the seed coat of H. tiliaceus seeds was nickedopposite to embryo slightly with nail cutter [5]. The filter paper was moistened with 10 ml of boric acid solutions containing 100 ppm, 150 ppm and 200 ppm boron respectively and kept in petridish.The filter papers moistened with 10 ml of distilled water served ascontrol. Experiments were carried out in the laboratory in the darkwith an average temperature ranging from 28-30 ± 2 °C. Germinationpercentage was recorded after every 24 hours and seedling growthwas studied after 120 hours. The seedlings were analyzed for Averageshoot length, Average root length and Average fresh weight. Vigourindex was calculated by using the formula proposed by Abdul-Bakiand Anderson, [6]. VI = (Root length + Shoot length) × germination percentage. The activity of enzyme Catalase (EC 1.11.1.6) was assayed following the method of Luck [7] as described by Sadasivam and Manikam [8] and expressed as Δ OD min-1 mg-1 protein. Peroxidase (EC 1.11.1.7) activity was studied by the method of Horiguchi, [9]. The enzyme activity was expressed as Δ OD h-1 mg-1 protein. The soluble proteins in the enzyme were determined according to method of Lowry et al., [10]. All the values presented in the table are due the replicate experiments.
Results and Discussion
Effect of Boron toxicity on seed germination
Influence of boron treatment on germination percentage of thethree Hibiscus species is depicted in Figure 1 and Table 1. It is clear from the Figure 1 that the germination percentage is decreased due to higher concentrations of boron in all the three species. About 80.00% (H. cannabinus), 97.82% (H. sabdariffa) and 13.33% (H. tiliaceus) of final germination of corresponding control is noticed under boron stress at 200 ppm level (Table 1). The glycophytic species H. cannabinus and H. sabdariffa showed relatively better tolerance to boron stress while, germination is markedly affected in halophytic species H. tiliaceus. Lower Boron levels (100 and 150 ppm) increased the germination percentage over control in H. sabdariffa.
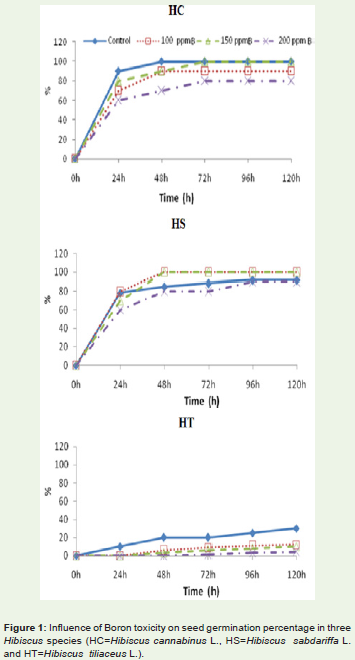
Figure 1: Influence of Boron toxicity on seed germination percentage in three Hibiscus species (HC=Hibiscus cannabinus L., HS=Hibiscus sabdariffa L.and HT=Hibiscus tiliaceus L.).
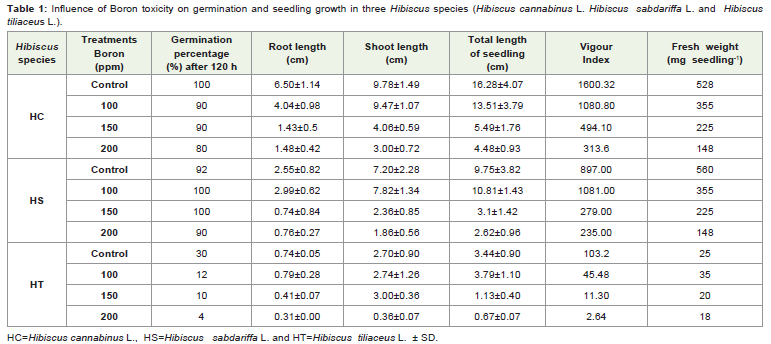
Table 1: Influence of Boron toxicity on germination and seedling growth in three Hibiscus species (Hibiscus cannabinus L. Hibiscus sabdariffa L. and Hibiscustiliaceus L.).
Two wheat (Wyalkatchem and Mace) and four barley (Hamelin,Mundah, Gardiner and Buloke) varieties for their tolerance todifferent boron (0, 5, 10, 25, 50, 100 and 150 mg B kg-1) concentrations and germination percentage was measured [3]. During early stages of germination, boron treatment showed significant differences between cultivars and concentrations. Boron concentration above 10 mg B kg-1 significantly reduced germination percentage during early stages of germination in wheat and barley cultivars. However, wheat cultivars are more tolerant than the barley cultivars at germination stage. Higher (150 mg B kg-1) boron concentration significantly reduced germination percentage in all the cultivars of wheat and barley on the first day performance of germination. He noticed inconsistent response of further germination in boron solution. He also stated that poorer germination may occur in cultivars sensitive to high boron treatments. In case of H. cannabinus and H. sabdariffa although the germination is delayed due to boron stress in initial phase the germination shows recovery latter and final germination percentage is only slightly lowered. But in case of H. tiliaceus inhibition of germination continues till the latter stages and it reflects sensitive nature of this species to boron stress.
Effect of Boron toxicity on seedling growth
The root length is decreased due to higher concentrations of boron in all the three Hibiscus species. About 22.77% (H. cannabinus),29.80% (H. sabdariffa) and 41.89% (H. tiliaceus) reduction of rootlength is noticed over control under boron stress at 200 ppm level. Thehalophytic species H. tiliaceus showed higher tolerance to boron stresswhile, glycophytic species H. cannabinus and H. sabdariffa showedlower tolerance, with respect to seedling root growth. But at the sametime slower growth of root in this species cannot be overlooked. Brdar-Jokanovic et al., [11] reported intercultivar differences regarding reduction in primary root length of wheat seedlings under ‘B’ stress. Such reduction was more prominent in cultivar Renan while, it was less in cultivar Trakija. Cultivar Stephens showed maximum primary root length among studied cultivars. According to Nable [12], tolerant wheat and barley cultivars maintain lower concentrations of boron in roots than sensitive genotypes. Liu et al., [13] observed that boron toxicity reduced the root cell division. This may be cause of reduction in root length due to ‘B’ toxicity. The shoot length is decreased due to 200 ppm concentrations of boron in all the three Hibiscus species. But at lower concentrations (100 ppm) in case of H. sabdariffa and (100 and 150 ppm) in case of H. tiliaceus there is increase in shoot length over control. In H. cannabinus there is continuous decrease in seedling shoot length along with increase in boron concentration. About 30.60 % (H. cannabinus), 25.83 % (H. sabdariffa) and 13.33% (H. tiliaceus) control of shoot length is noticed under boron stress at 200 ppm level. The glycophytic species H. cannabinus showed higher tolerance to boron stress while, halophytic species H. tiliaceus showed lower tolerance, with respect to seedling shoot length at this boron concentration. According to Nable [12], tolerant wheat and barley cultivars maintain lower concentrations of boron in shoots than sensitive genotypes. Boron toxicity symptoms have been quoted to start occurring at 14 mg B kg-1, for barley [14]. The shoot lengths of seedlings of the two chickpea cultivars (Go¨kce and Ku¨smen) were affected differently due to B treatment. Increasing concentrations of B (1.6 and 6.4 mM) significantly increased shoot length in cultivar Go¨kce, but decreased shoot length of cultivar Ku¨smen [15]. These workers found higher ‘B’ accumulation in cultivar Ku¨smen thanin cultivar Go¨kce and it has been demonstrated that ‘B’ toxicity iscorrelated with intracellular ‘B’ concentration [16]. Boron at 10 mg B kg-1 caused greatest mean shoot length, suggesting need of boron during early growth [3]. Such stimulation of shoot growth is also seen in H. sabdariffa and H. tiliaceus at low ‘B’ concentration. The tolerance mechanisms amongst plants are very variable, however the tolerance mechanism to reduce uptake of boron in shoot seems to be common as that of roots. According to Farr [3], shoots show significant reduction in size and growth under boron toxic conditions. At 200 ppm ‘B’ levels similar picture is seen in all the three Hibiscus species. Boron treatment might be influencing root and shoot growth in seedlings of these three species in a different manner. In case of H. cannabinus there is greater decrease in root length as compared to the shoot length. In case of H. sabdariffa there is greater decrease in shoot length due to high ‘B’ concentration. In H. tiliaceus there is no consistent effect of ‘B’ on root to shoot growth.
The seedling length is decreased due to higher concentrations ofboron in all the three species. It is further evident from Table 1 that, the seedling length in all the three species decreased with increasing boron concentration. Except at lower concentrations (100 ppm) H. sabdariffa and H. tiliaceus exhibited higher seedling length. About 27.52% (H. cannabinus), 26.87% (H. sabdariffa) and 14.83% (H. tiliaceus) of seedling length of control seedlings is noticed under boron stress at 200 ppm level. The glycophytic species H. cannabinus and H. sabdariffa showed approximately equal tolerance to boron stress while, halophytic species H. tiliaceus showed less tolerance with respect to seedling length. The vigour index is decreased continuously along with increase in concentrations of boron in H. cannabinus and H. tiliaceus. In case of H. sabdariffa vigour index is slightly increased at 100 ppm ‘B’ but thereafter there is marked decline. It is further evident from Table 1 that, the vigour index in all the three species is considerably decreased at highest (200 ppm) boron concentration except at lower concentrations (100 ppm) H. sabdariffa showed higher value over control. About 19.59% (H. cannabinus), 26.19% (H. sabdariffa) and 2.55% (H. tiliaceus) of vigour index is noticed over control under boron stress at 200 ppm level. The glycophytic species H. sabdariffa showed higher tolerance to boron stress while, halophytic species H. tiliaceus showed lower tolerance, with respect to vigour index. Although there are no reports are available about influence of boron toxicity on vigour index of seedlings, the observations of Paull [14] indicated that wheat plants subjected to boron toxicity showed low vigour. Influence of boron toxicity on fresh weight of three Hibiscus species is depicted in Table 1. It is evident from the Table 1 that the fresh weight per seedling is decreased due to higherconcentrations of boron in H. cannabinus and H. sabdariffa. In H.tiliaceus the fresh weight of seedlings treated with 100 ppm B is slightly higher but there after there is decline in fresh weight. The decrease in fresh weight (in comparison to control) at highest concentration (200 ppm) is more significant in H. cannabinus and H. sabdariffa as compared to H. tiliaceus. At seedling stage uptake of water mainly contributes to fresh weight besides increase in cell number and cell size. It is obvious that boron toxicity affects all these three parameters in the three Hibiscus species. About 28.03% (H. cannabinus), 26.43% (H. sabdariffa) and 72.00% (H. tiliaceus) of control fresh weight is noticed under boron stress at 200 ppm level.
Effect of Boron toxicity on antioxidative enzymes
Effect of boron (B) toxicity on activity of enzyme catalase (CAT)and peroxidase (POX) in seedlings (120 h) of H. cannabinus, H.sabdariffa and H. tiliaceus is shown in table 2. It is clear from Table 2, that the maximum enhancement of CAT and POX activity in case of H. sabdariffa seedlings is noticed under high boron concentrations 150 ppm and 200 ppm respectively. A slight stimulation of CAT activity is also visible in case of H. cannabinus and H. tiliaceus, due to treatment of 100 ppm concentration of boron. The activity of enzyme POX is declined in seedlings of H. cannabinus and H. tiliaceus due to entire range of boron treatment. Boron toxicity is reported to lead to synthesis and accumulation of ROS in case of Citrus leaves [17], barley leaves [18] and apple rootstocks [19]. Antioxidant enzyme catalase plays important role in detoxification of H2O2 generated during dismutation of superoxide radical by SOD along with oxygen [18]. Dube et al., [20] reported that excess boron in sunflower leaves caused increase in catalase activity. There are several reports which indicate that the boron toxicity brings about increase in catalaseactivity in tobacco [21], barley [22] and in grapevine [23]. Similarly,apple rootstocks [24] and tomato [25] exhibited increased activity of catalase in response to B toxicity. In contrast to above reports, there are few reports of reduction in CAT activity in response to boron treatment in plants. Roots have lower catalase activity than leaves in wheat and barley under boron stress [3]. Keles et al., [17] also reported decreased activity of CAT in citrus leaves due to B toxicity.
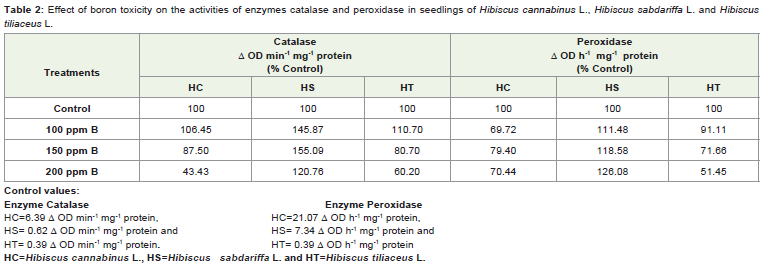
Table 2: Effect of boron toxicity on the activities of enzymes catalase and peroxidase in seedlings of Hibiscus cannabinus L., Hibiscus sabdariffa L. and Hibiscustiliaceus L.
In the leaf tissue of C3 species such as tobacco, sunflower,barley, tomato and chickpea due to photorespiratory process thereis accumulation of H2O2 and catalase localized in peroxisomes plays important role in breakdown of H2O2. Hence, increase in catalase activity in the leaf tissue may takes place to regulate H2O2 level elevated as a result of increased photorespiration in response to boron treatment. An increase in SOD activity in roots and shoots in response to boron stress can also result in elevation of H2O2 level [15,26]. In seedling tissue probably due to disturbances in respiratory electron transport ROS production can take place under boron toxicity and the increase in catalase activity may help in scavenging the H2O2 level. Among the three Hibiscus species, seedlings of H. sabdariffa possess such adaptive feature as the activity of catalase is markedly increased due to boron treatment. In the other two species however, such stimulation in catalase activity is noticed only in response to lower dose (100 ppm) of boron.
Peroxidase is an oxidative enzyme which plays multiple roles in thecells. The enzyme play important role in secondary metabolism andlignin biosynthesis and possible involvement of this enzyme in auxinmetabolism has been also discussed. Peroxidase plays important rolein detoxification of H2O2 generated during dismutation of superoxide radical by SOD along with oxygen [18]. This is due to the fact that there are several isozymes of peroxidase which are localized in different regions and organelles in the cell. In view of Castillo, [27] an increase in POX activity may be an early response to several stress like B and may provide cells with tolerance to H2O2 formed when plants are exposed to stress. Palavan-Unsal et al., [28] reported promotion of activity of enzyme POX under ‘B’ stress and also detected presence of POX in lignified tissues.
POX activity was found to be increased in shoots of twochickpea cultivars due to 6.4 mM B treatment [15]. Above reports on the behavior of enzyme peroxidase are from full grown plants and germinating seeds are seldom investigated. Bhamburdekar [29] noticed a decline in peroxidase activity in pigeonpea seedlings due to boron toxicity. Such decline in enzyme activity is evident in seedlings of two Hibiscus species H. cannabinus and H. tiliaceus in response to boron toxicity but in case of H. sabdariffa opposite trend is noticed. This free radical scavenging enzyme system appears to be more efficient in H. sabdariffa than H. cannabinus and H. tiliaceus under the condition of boron excess.
Conclusion
From foregoing account of result it is concluded that H. sabdariffais more tolerant to boron stress than H. tiliaceus and H. cannabinuswith respect germination percentage, seedling growth, vigour indexand activities of antioxidative enzymes. An interspecific differencein boron tolerance at germination stage was evident in the threeHibiscus species.
Acknowledgement
One of the authors, B. A. Sonar is gratefully acknowledges UGCfor the award of fellowship for Meritorious Students in Science.Authors are thankful to The Head, Department of Botany, ShivajiUniversity, Kolhapur for availing the necessary laboratory facilities.
Conflict of Interest
Authors declare that they have no conflict of interest.
References
- Nable RO, Bañuelos GS, Paull JG (1997) Boron toxicity. Plant Soil 193: 181-198.
- Yau SK, Ryan J (2008) Boron toxicity tolerance in crops: a viable alternative to soil amelioration. Crop Sci 48: 854-865.
- Farr HJ (2010) Early growth tolerance to boron and salt in wheat and barley. A dissertation is submitted as a part of the requirement for the award of the Degree of Bachelor of gribusiness with Honours (Agricultural Technology) at Curtin University.
- Wimmer MA, Mühling KH, Läuchli A, Brown PH, Goldbach HE (2003) The interaction between salinity and boron toxicity affects the subcellular distribution of ions and proteins in wheat leaves. Plant Cell Environ 26: 1267-1274.
- Elevitch CR, Thomson LAJ (2006) Hibiscus tiliaceus (Beach hibiscus) Malvaceae (mallow family) Species Profiles for Pacific Island Agro-forestry.
- Abdul-Baki AA, Andersion JD (1973) Vigor determination of soybean by multiple criteria. Crop Sci: 13: 630-633.
- Luck H (1974) In: Methods in Enzymatic Analysis II (ed.) Bergmeyer. (Publ.) Academic Press, New York; pp. 885.
- Sadasivam S, Manikam A (1992) Biochemical methods for agricultural sciences. (Publ.) Wiley Eastern Ltd. New Delhi: 105pp.
- Horiguchi T (1988) Mechanism of manganese toxicity and tolerance of plants IV. Effects of silicon on alleviation of manganese toxicity of rice plants. Soil Sci Plant Nutr 34: 65-73.
- Lowry OH, Rosenbrough NJ, Furr AL, Randall RJ (1951) Protein measurement with folin phenol reagent. J Biol Chem 193: 265-275.
- Brdar-Jokanović M, Maksimović I, Nikolić-Äorić E, Kraljević-Balalić M, Kobiljski B (2010) Selection criterion to assess wheat boron tolerance at seedling stage, primary vs. total root length. Pak J Bot 42: 3939-3947.
- Nable RO (1988) Resistance to boron toxicity amongst several barley and wheat cultivars: A preliminary examination of the resistance mechanism. Plant Soil 112: 45-52.
- Liu D, Jiang W, Zhang L, Li L (2000) Effects of boron ions on root growth and cell division of broadbean (Vicia faba L.). Israel J Plant Sci 48: 47-51.
- Paull JG (1990) Genetic studies of the tolerance of wheat to high concentrations of boron. Doctor of Philosophy Thesis, University of Adelaide.
- Ardic M, Sekmen AH, Tokur S, Ozdemir F, Turkan I (2009) Antioxidant responses of chickpea plants subjected to boron toxicity. Plant Biol 11: 328-338.
- Reid RJ, Hayes JE, Post A, Stangoulis JC, Graham RD (2004) A critical analyses of the causes of boron toxicity in plants. Plant Cell Environ 25: 1405-1414.
- Keles Y, Oncel I, Yenice N (2004) Relationship between boron content and antioxidant compounds in Citrus leaves taken from fields with different water sources. Plant and Soil 265: 343-353.
- Zhu Z, Wei G, Li J, Qian Q, Yu J (2004) Silicon alleviates salt stress and creases antioxidant enzymes in leaves of salt stressed cucumber (Cucumis sativus L.). Plant Science 167: 527-533.
- Molassiotis A, Sotiropoulos T, Tanou G, Diamantidis G, Therios I (2006) Boron induced oxidative damage and antioxidant and nucleolytic responses in shoot tips culture of the apple rootstock EM 9 (Malus domestica Borkh). Environ Exp Bot 55: 54-62.
- Dube BK, Sinha P, Chatterjee C (2000) Boron stress affects metabolism and seed quality of sunflower. Trop Agric 77: 89-92.
- Garcia POC, Rivero RM, Lopez-Lefebre LR, Sanchez E, Ruiz JM, et al. (2001) Response of oxidative metabolism to the application of carbendazim plus boron in tobacco. Aust J Plant Physiol 28: 801-806.
- Karabal E, Yucel M, Oktem HA (2003) Antioxidant responses of tolerant and sensitive barley cultivars to boron toxicity. Plant Sci 164: 925-933.
- Gunes A, Soylemezoglu G, Inal A, Bagci EG, Coban S, et al. (2006) Antioxidant and stomatal response of grapevine (Vitis vinifera L.) to boron toxicity. Sci Hort 110: 279-284.
- Sotiropoulos TE, Molassiotis A, Almaliotis D, Mouhtaridou G, Dimassi K, et al. (2006) Growth, nutritional status, chlorophyll content, and antioxidant response of the apple rootstock MM 111 shoots cultured under high boron concentrations in vitro. J Plant Nutr 29: 575-583.
- Cervilla LM, Blasco B, Rıos J, Romero L, Ruiz J (2007) Oxidative stress and antioxidants in tomato (Solanum lycopericum) plants subjected to boron toxicity. Ann Bot 100: 747-756.
- Ardic M, Sekmen AH, Turkan I, Tokur S, Ozdemir F (2009) The effects of born toxicity on root antioxidant systems of two chickpea (Cicer arietinum L.) cultivars. Plant Soil 314: 99-108.
- Castillo FJ (1992) Peroxidases and Stress. In: Plant Peroxidases 1980-1990. Topics and Detailed Literature on Molecular, Biochemical and Physiological Aspects, (Eds) Penel C, Gaspar TH, Greppin H. University of Geneva Press, Geneva. 187-203.
- Palavan-Unsal NC, etin E, Kadıoglu A (2000) Boron stress affects peroxidase activity. Plant Peroxidase Newsletter 15: 37-44.
- Bhamburdekar SB (2002) Germination studies in pigeonpea (Cajanus cajan L.). PhD Thesis, Shivaji University, Kolhapur.