Research Article
Symptomless Transmission of Seed Borne Botrytis Cinerea into the Seed of Next Generation Lettuce Plant (Lactuca Sativa)
Yahaya SM1*, Fagwalawa LD1, Ali MU1, Umma M1, Lawan M1, Bilkisu A1 and Amina LA2
1Department of Biology, Kano University of Sciences and Technology, Wudil P.M.B. 3244, Kano State Nigeria
2Department of Biochemistry, Kano University of Science and Technology Wudil P. M.B.3244. Kano State-Nigeria
Corresponding author: Yahaya SM, Department of Biology, Kano University of Sciences and Technology, Wudil -Kano State P.M.B. 3244, Nigeria; E-mail: sanimyahya@gmail.com
Citation: Yahaya SM, Fagwalawa LD, Ali MU, Abdullahi H, Umma M, et al. Symptomless Transmission of Seed Borne Botrytis Cinereainto the Seed Of Next Generation Lettuce Plant (Lactuca Sativa). J Plant Sci Res. 2016;3(1): 140.
Copyright © 2016 Yahaya SM, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Plant Science & Research | ISSN: 2349-2805 | Volume: 3, Issue: 1
Submission: 11/12/2015; Accepted: 04/01/2016; Published: 16/01/2016
Abstract
Study was carried out to determine the behaviour of endophytic and systemic pathogen Botrytis cinerea the causative agents of grey mould disease in ornamental and vegetables. The necrotic pathogen was isolated from Botrytis selective media from sterilised portion of lettuce seeds collected from the plants which were dry inoculated at seedlings and flower stage of development. Seed collected from first-experiment plants, which was found to have carry the B. cinerea inoculum used during dry inoculation, was used to grow plants for the second generation experiment at controlled environment room after surface sterilization. Isolation of the inoculating isolate from the seed of healthy second generation lettuce plants confirmed symptomless transmission of the inoculating isolates.
Keywords: Botrytis cinerea; Endophytic infection; Systemic infection; Sterilization; Lactuca sativa
Introduction
Botrytiscinerea (teleomorph Botryotinia fuckeliana) have beenrecognized as the most important pathogen affecting nursery plants,vegetables, ornamental, field orchard and forestry plants as well asstored and transported agricultural products throughout the world[1]. At the early times of its recognition, it was referred as a temperatearea pathogen, nevertheless currently the pathogen’s impact has beena concern in all parts of the world ranging from tropical, sub-tropicalto world cold areas. The pathogen produces clear grey conidia onLong branch conidiophores [2], the conidia infect the plant directlyto cause spreading necrotic lesions. In some plant B. cinerea can enterand remain dormant or quiescent for varying period of time [3].During quiescence further fungal growth and colonization are halted, and sign of the pathogen is not visible but became activated by theonset of senescence or stress of the host tissue [4,5].
In a previous research [6,7] showed that fungal diseases aremainly transmitted via seed, and seed borne infection can lead tosymptomless infections after they have been transmitted to seedlings.They reported that B. cinerea can grow into the plant without showingsymptoms and single isolates may grow throughout the healthy plantwithout showing symptoms. They reported that the infections appearto be seed-borne, but may also arise from air-borne spores. Study by[8] who isolated Botrytis fabae, the cause of chocolate spot of Vicia fabafrom bean seeds found that seed infection does not always transmitinfection into seedling, [8] reported that the levels of B. fabae conidiaon most infected seeds, which were tested, were too low to cause an aggressive lesion at 150C and the fungus dies without seriouslydamaging the plant. He concluded that for an aggressive infection tooccur, seeds would have to carry a high amount of inoculum. This wassupported by [9] who reported that not all infections by endophyticB. cinerea results in systemic infection. The evidence was (a) failureto detect B. cinerea from sections of surface sterilized asymptomaticepicotyl from seedlings with visible root lesions and (b) infected seedsgave rise to apparently looking healthy seedlings.
In this experiment, B. cinerea was isolated from seeds as well asleaves of symptomless lettuce plants. Similar, report by [6,10]. Theyfound that seed infection lead to systemic endophytic infection evenwhen the external inoculum was excluded. They concluded that theinfection was internal since B. cinerea was isolated from the surfacesterilised seeds of Primular and lettuce plants. Therefore, the presentstudy tested two hypotheses. First, the B. cinerea grows systemicallyin lettuce plant and second, the infection from infected host seedlingcould be transferred to the seed in lettuce that subsequently infectseeds of next-generation plant without showing symptom.
Material and Methods
Seed Sterilization
Lettuce seed (var. Tom Thumb), purchased from FothergillsSeeds, New market, and was used for the experiments. Portion of theseed was sterilized in fungicide by soaking in100ml of (0.1g/l) systemicfungicide Shirlan (active ingredient 500g/l Fluazinam. Sygenta CropProtection UK limited) for 2h and drying over night before sowing.
Inoculation and Seedling growth
Seeds were sown in16 trays filled with vermiculite growingmedium. These trays were kept in a glass house at University ofReading. Eight trays were used each for sterilized and non-sterilizedseeds. One week after germination, four trays of seedlings growingfrom sterilized and non- sterilized seeds (Figure 1) were selected andcovered with black polyethylene bags. Botrytis cinerea spores 2.20incubated for two weeks were used for the inoculation. Dry sporeswere harvested from the petri plates by tapping gently on a sterilepiece of aluminium foil. The spores were transferred into 10ml sterilesyringes (BD Plastipak, UK). The syringes were fitted with 25mm63/100 23GX1 needles. The needles were inserted into the bag beforeforcibly delivering the spores into the enclosed area, and left for24h, to facilitate germination of spores through building up of highrelative humidity. To estimate the number of spores deposited perunit area an acetate paper on which graph sheet was photocopied wasplaced in each tray and examined by a microscope. There was averageof 12 spores/ mm.2.A week after seedling inoculation seedlings weretransplanted in to15cm pot and placed in 20 blocks of 4 treatments(Figure 2).

Figure 1: Dry spore inoculation of seedling.

Figure 2: Dry spore inoculation of seedling.
Dry spore inoculation of the flower/bud
At flower stage 10 blocks consisting of 40 plants (half of thereplicates,) were selected from the first four treatments (a) fungicidetreated seed/uninoculated seedling (b) fungicide treated seeds/inoculated seedlings (c) non fungicide treated seeds/ uninoculatedseedlings (d) non fungicide treated seeds/ inoculated seedlings.Flower and bud of the plants were covered with black polyethylene bags and inoculated with similar spores as in seedling inoculations,and left for 24h.
Isolation of Botrytis cinerea from seeds of lettuce plants
To determine the systemic transmission of infection into theseed, 10 seeds from each of the plants grown in the eight treatments oflettuce plants were collected and sterilized by soaking in 20ml of 70%ethanol in a duram bottle for 30 minutes, and allowed to dry for onehour in the laminar flow. The seeds from each plant were separatelycrushed and plated on Botrytis selective media and incubated at180C for one week. Growth of Botrytis cinerea from each seed wasmonitored, counted and recorded.
Genetic characterization of B. cinerea isolates
Using pestle and mortar in the presence of liquid nitrogen, 100mgmycelium of the seed isolates grown in MEA plates was scratched andground to fine powder. DNA was extracted from the fine powderusing DNeasy Plant Mini Kits (Qiagen, West Sussex, UK); theconcentration of the DNA was adjusted to 10ng μl -1 using NanodropND 1000 Spectrophotometer and stored at -20 °C until needed.
The template DNA was run in polymerase chain reactions with scarprimers designed by [11] which are specific for B. cinerea. Templatesthat gave DNA band were characterized by nine microsatellitemarkers published by [12] labelled with FAM (Blue), HEX (Green),and NED (Yellow). The volume for each reaction mixture contained25 μl of Biomix (Bioline, UK), 20 μl of water, 1.5 μl of each forward and reverse primers and 2 μl of DNA template. The PCR of theSSR was repeated several times for successful amplification. Theproducts from successful amplifications were multiplexed in threecombinations; (BC1, BC4, BC9), (BC3, BC6, BC10), (BC2, BC5, BC7).These groups were chosen to avoid overlapping the allele size rangesof the primers in each mix. The PCR was run at initial denaturingstep of two minutes at 94 °C for BC1, BC2, BC3, BC5, BC6 and BC9followed by 35 cycles of one minute at 94 °C, one minute annealingat 53 °C, 30 seconds at 72 °C and 5 minutes at 72 °C. While BC4, BC7and BC10 have an annealing temperature of 590C. The products weremultiplex submitted for fragment analysis at Macrogen South Korea.The results from the fragment analysis were interpreted by usingGenemapper V4 version software (Applied Biosystem).
Transmission of infection from seed to the next generationplant
To test whether B. cinerea has been successfully transferred fromseed to the healthy next plant generation seed, infected seeds werecollected from five flower inoculated plants D2-2, D2-3, D2-5, D2-10and A2-6. The seed from each plant was divided into two. One-halfof the seed was sterilized with fungicide as previously. Sterilized andnon-sterilized seeds from all the five plants were grown in 10, 20mmpots. Five plants were grown for each of the two treatments, one plantper pot. The plants were grown up to seedling stage (four month),with seed’s collection from all the plants (Figure 3 and 4).

Figure 3: Second generation lettuce plant at seedling stage.

Figure 4: Second generation lettuce plant grown at Control environment atFlower stage.
Experimental design
First generation experiment-Glasshouse. Seedling to seedtransmission of B. cinerea. Prior to sowing seeds, 100 seeds from bothsterilized and non-sterilized were plated in BSM plate to determine the initial level of infection. Fifty seeds from each of the two treatmentswere crushed before plating. Plates were incubated at 180C for 1 week.
The experiment design used a factorial block design in whichall the treatments were assigned to each block. The main factors(a) fungicide treatment of the seed, (b) seedling inoculation, (c)flower inoculation. At two leaf stage, seedlings were dry inoculatedwith spores of isolate 2.20. A week after inoculation seedling weretransplanted into 80, 15 cm pots and arranged in randomized blocks.Two benches were used all in a single glasshouse (twenty blocks offour treatments). Each block has one plant from each of the initialfour treatments: (a) seed sterilization/uninoculated seedling, (b)sterilized seed/inoculated seedling, (c) non fungicide treated seed/uninoculated seedling, (d) non sterilized seed/ inoculated seedling.When the plants were two month old using 15mm cork borer leavessamples was collected from all the 80 plants and plated on BSM. Atflower stage, plants were divided into two portions each consistingof 10 blocks of 40 plants. Flower and bud of the second portion wascovered with black polyethylene bag and dry inoculated with sporeisolates of B. cinerea 2.20 as in seedling inoculation. Matured seedswere collected from all the plants.
The Second generation experiment transmission of B.cinerea wasfrom first generation to second generation plants without showingsymptoms. To determine whether B. cinerea can be transmitted fromseed to the plant without showing symptom, infected seed from flowerinoculated plants were grown in a controlled environment which hasfewer chances of external contaminations. The experimental designwas randomized block design with seed sterilization as the mainfactor. Each block has two replicates. The two treatments were grownup to seedling stage. Matured seeds were collected from all the plantsand genetically characterized. Peak height of the microsatellite datawas converted into scores of one and zeros. One and zeros representedthe presence or absence of bands of a given size respectively.
Statistical analysis
Significance of treatment effects on incidence of seed infectionby B. cinerea was determined by factorial analysis of variance. Asignificant value in the ANOVA indicates a significant relationship inthe flower inoculation P< 0.05.
Results
Seed infection
One hundred seeds of Lettuce var. Tom Thumb were placed onBotrytis selective media but no infection was detected. Infection wasalso not recorded on 100 sterilized seeds of this variety before plating.However, infection was detected when both fungicides treated anduntreated seeds were crushed before planting on Botrytis selectivemedia.
Systemic infection of B. cinerea in lettuce leaves
Botrytis cinerea was detected from leaves of plants grown fromsterilized and non-sterilized seeds at Glass house. Infection wasdetected from the two treatments. More than half of the plants carriedleaf infection of B. cinerea, the incidence of B. cinerea in leaf of plantsgrown from sterilized seeds [22% (9/40)] was similar to leaf of plants grown from non-sterilized seeds [30%(12/40)] but not significantlyso. While detection of B. Cinerea from new lettuce leaves in seedlingsgrown from sterilized seed and inoculated with fungal spores at 2 leafstage shows that the proportion of infected leaves was the same inplants grew from treated and untreated seed. But inoculated plantshad a higher incidence of leaf infection. This was unaffected by seedsterilization (Table 1).
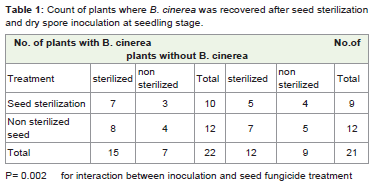
Table 1: Count of plants where B. cinerea was recovered after seed sterilizationand dry spore inoculation at seedling stage.
Transmission of B. cinerea from lettuce Seedling to the seedin glass house
Out of the seed plated on BSM plate a total of 364 seeds a crossthe eight lettuce treatment shows infection ofBotrytis cinerea. Seedinfection in plants grown from sterilized seed was 32% (117/364),while in seeds collected from plants inoculated at seedlings stage was27% (100/365), while infection in seeds collected from inoculatedat flower/buds stage was 41 % (148/364). More infected seeds wererecorded from plant inoculated at flower/buds stage irrespectiveof seed sterilization and seedling inoculation F 1, 79 =23.71, P< 0.001 (Table 2). The effects of flower/bud inoculation and seedlinginoculation were independent of one another on the seed infection byB. cinerea F 1,79=0.60P = 0.441(Table 2,3,4).

Table 2: Effects of flower/bud inoculations and seedling inoculation seedinfection by B. cinerea
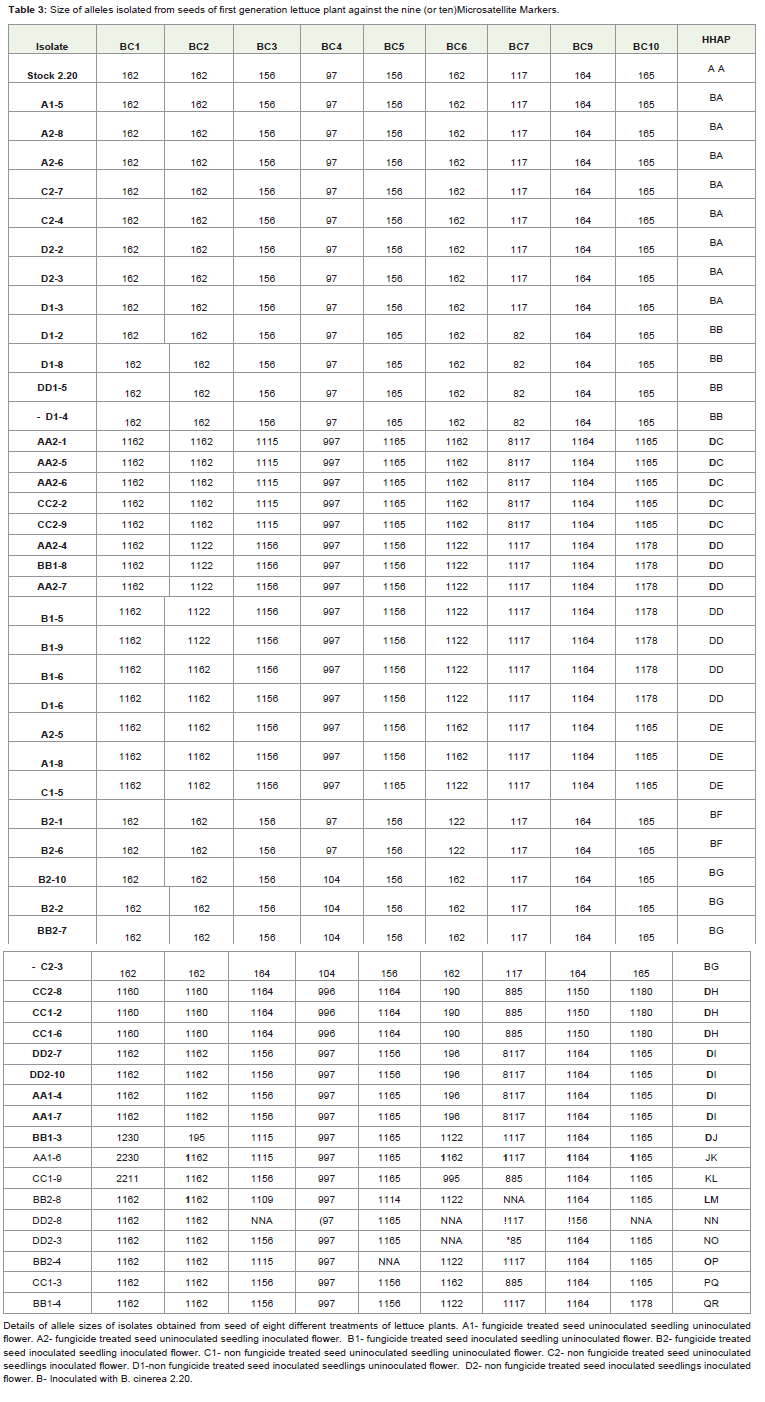
Table 3: Size of alleles isolated from seeds of first generation lettuce plant against the nine (or ten)Microsatellite Markers.
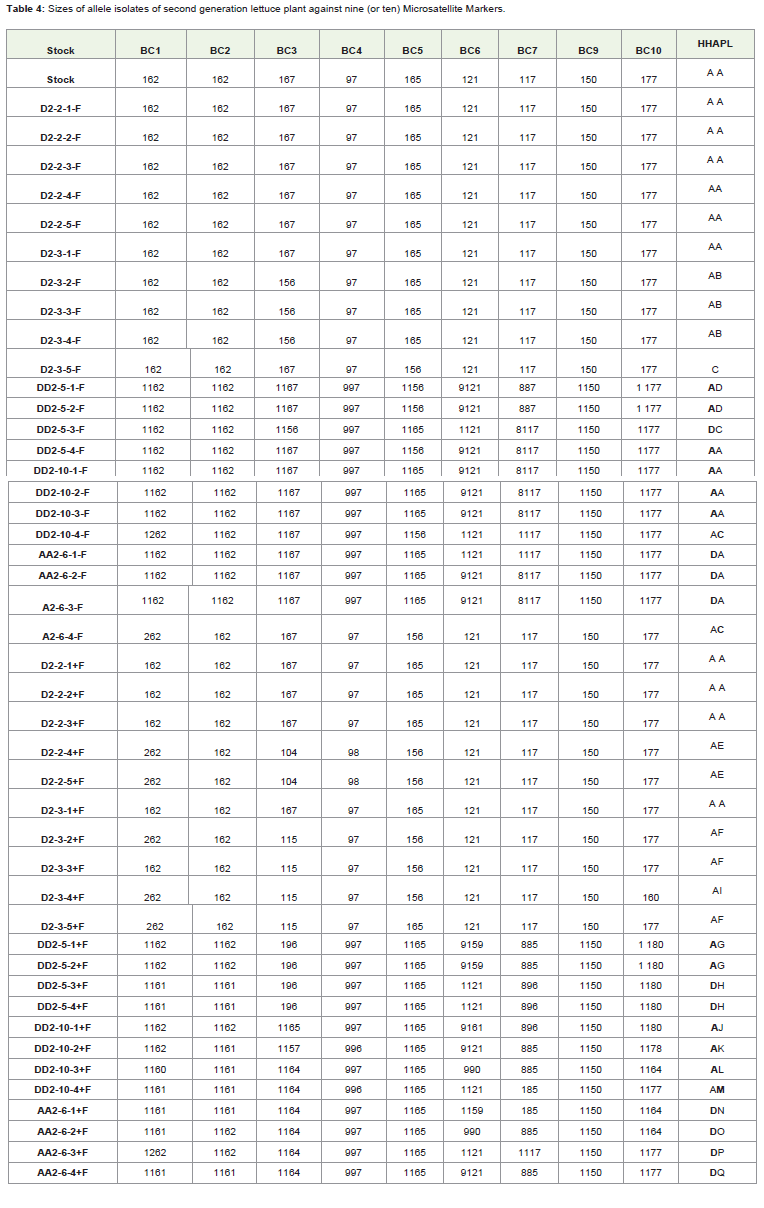
Table 4: Sizes of allele isolates of second generation lettuce plant against nine (or ten) Microsatellite Markers.
Microsatellite of seed isolates
Isolates recovered from seed of the glass house grown lettuceplant were confirmed as B. cinerea isolates by polymerase chainreaction using scar primers designed by [13]. The isolates were thengenotyped using nine microsatellite markers published by [13]. Theisolates used to artificially inoculates seedlings, 2.20 (coded haplotypeA), was recovered from some of the plants, whether inoculated ornot. However, the length of the isolates detected with microsatelliteloci BC1 and BC3 were found to be lower than what was obtained bythe previous workers (result not shown). The microsatellite primers separated isolates into 17 different haplotypes. The frequency ofhaplotype’s existence within the populations is depicted in Figure 1. One haplotype (47%) was recovered from seed of few flowerinoculated plants and seed of some non flower inoculated plantsresult not shown.
Out of the isolates collected from seed of plants grown fromsterilized seed 8.3% of the haplotypes matches the isolate 2.20 usedin the inoculation, while 91% of the isolates were different from theinoculated isolates. While in seed collected from seedling inoculatedplant, 15% of the haplotypes detected genetically matches isolate usedfor inoculating the plants, while 85% were different.
Overall, the most commonly occurring haplotype whichdominated isolates collected from the seed of flower inoculated plants(22%) was similar to the isolate used for seed and flower inoculationat the first experiment. However, the inoculating isolate was notdominant in uninoculated plants (9%).
Transmission of infection from Seed of first generationplants to the healthy next generation plant
Seed infection was recovered from all the second generationlettuce plant grown at controlled environment room. Out of theisolates recovered from seed of plant grown from untreated seeds59.1% of the isolated resemble the inoculating isolate, while 40.9%differ from the isolate used in the inoculation of first-generationplants. However, all the seed isolates recovered from plants grownfrom sterilized seeds have different genotypic identification with theisolates used in the inoculation of first-generation plants. Surprisingly,like isolates of the first generation plants, the length of the genotypeobtained with microsatellite loci BC1 and BC3 were lower than thelength reportedby the previous workers.
Discussion
The study found that first healthy lettuce plants without anyvisible symptoms may harbour B. cinerea in leaves, stems, root andseeds, second that as the plant growsB. cinerea infection initially inlettuce seeds or seedlings spread to the seed. Third seed infectiontransmitted into the next seed generation is a complete systemic andendophytic infection. Systemic, symptomless progression has alsobeen demonstrated earlier in lettuce apart from commercial hybridPrimula [6,7].
In 1992 Singh [14] reported positive evidence of transmission ofBotryodiplodia theobromae from maize seed to seedlings. Seeds whichwere heavily infected failed to germinate and were covered withprofuse growth of pycnidia on their surfaces. However, those withweak to moderate inoculum of Botryodplodia theobromae germinatedand their histopathology showed that the fungus readily invadesgrowing seedling from adjoining pericarp, scutellum or closing tissue.They concluded that the fungus grows internally in the seedlings.Richard et al. showed that Aspergillus flavus become systemic inyoung maize seedlings grown from contaminated seed. It was shownthat germination rates were considerably lower when Aspergillusflavus entered and contaminated the seed. The distribution of thefungus suggested that initially the organism may follow the meristemof the plant.
The present study established that artificial inoculation of plantswith B. cinereaallowed moderate re-isolation of the same isolate fromthe seed collected from the plants despite the fact that seeds fromall the plants were sampled for re-isolation. However, B. cinerea2.20 used for inoculation was recovered from the seeds of 22% offirst-generation plants. This indicates that inoculation and naturalinfestation from air borne spores and /or seed infection results ina successful endophytic establishment and systemic spread of B.cinerea in the glasshouse grown lettuce plant. B. cinerea 2.20, usedin the seedling and flower/bud inoculation was recovered from theseed of some flower, and few seedlings inoculated plants. Thesefindings agree with the results of [12] who reported moderate geneticvariation within B. cinerea isolates collected from fig plants in the same location in California but no differences between differenthosts. Similarly [15] found that B. cinerea populations from differenthost’s species (Strawberry, dandelion, black current, primula)sampled from different locations of the UK were genetically variableand most importantly highly variable within hosts. Nevertheless, inthe study conducted [15] all the isolates recovered from inoculatedand uninoculated lettuce plants were identical to the isolate used forinoculation. This indicates that a successful artificial inoculation athigh concentrations can eliminate natural infestations and spreadwithin plant populations. Mixed genotype infections are common inthe plant pathogen system; however, their impact on the course of theinfection and especially on the pathogen virulence and host responseto infection are poorly understood.
The study also establishes that inoculation of the flower/ bud madea significant difference to the infection levels with seed collected fromflower/bud inoculated plant having more seed infection (P=0.001)than seed collected from non flower/bud inoculated plant, so flower/bud inoculation has been successful (SE=0.452). The higher infectionof seed collected from flower/bud inoculated plant could be due tothe growth stage at which the plants were inoculated. The evidencehere are many studies have shown that flower is the major point ofentry of infection into the plant. Infection has also been detected inpart of crop inflorescence i.e., stylar infection in grapes [16-18]. Thefact that B. cinerea has been frequently isolated from various partsof crop inflorescence, it may probably be the most common site ofentry into the seed and other part of the plants. There is a need toinvestigate further the growth stage and infection in flower inoculatedplant. However, [14] found both seedling and flower inoculated plantto have more seed infection than uninoculated ones. Ochoa and Ellis[18] found that Fusarium oxysporum was seed-borne in commonnaranjilla (Solanum quitoense) and can be transmitted from seed toseedling systemically. Fusarium oxysporum was isolated from seedsand plant parts, which were surface sterilised before plating. Also[19,20] isolated several fungi from onion seeds and seedlings, whichwere seed-borne, but only Aspergillus niger and Fusarium oxysporumwere transmitted from seed to onion sets.
Plant grown from sterilized seeds/uninoculated seedling/flower/bud inoculations, had the highest seed infection, followed byseed collected from fungicide treated plants/inoculated seedlings/inoculated flower. This indicates that seed fungicide treatmenthas no effect on seedling or flower/bud inoculations. Least seedinfection was recorded in plants grown from non fungicide treatedseed/uninoculated seedling and uninoculated flower/bud. The factthat seeds collected from plant grown from non sterilized seed withneither seedling, nor flower/bud inoculations gave rise to infectedseeds means that the inoculum is either from the seed or enter fromsomewhere in the glasshouse. These results suggest that B. cinereacould move from one part of a plant to another implying that it isnot localised.
A moderate level of genetic diversity was found among isolaterecovered from seed collected from plant grown from fungicidetreated seeds. It is possible the fungicide treatment eliminated seedborne infection, but the plant can be more frequently attachedby airborne inoculum. More importantly, seed collected fromplant grown from non fungicide treated plants carried seed borneinfection, which systemically spread into the plants and the frequencyof these plants being attached by the air borne inoculum was less.This partly supports the result of [7]. They found that 56 B. cinereaisolate from different glasshouses in South Carolina (USA) wereall genetically different but found some clustering in relation tofungicide sensitivity. They suggested that B. cinerea populationswere genetically very diverse even in relation to fungicide resistance.However, no fungicide resistance test was carried out in the presentresearch. As reported [13] evidence of systemic infection spreadinginto plants from seed borne inoculum in lettuce plant. He discoveredthat isolates from leaves, stem and roots of plants were geneticallysimilar or independent of the various parts. Results of the presentstudy show that re-isolation of the inoculated isolated was successful, which provides evidence of systemic infection.
It was interesting to observe that one of the haplotypes occurredin high frequency among the isolate collected from seed of plantsgrown from non fungicide treated seed at second generationexperiment. Although few air borne conidia may exist in thecontrolled environment room, clearly the isolates originated from theseed infection.
However, the result of the fragment analysis was somewhatdifferent; the results of microsatellite loci BC1and BC3 at firstgeneration were found to be lower than the length obtained byprevious workers [14,20]. Furthermore, the abnormality was againnoticed from the results of second generation experiments for the sameprimers despite the fact that a new set of microsatellite primers wereused. The differences might be due to mutation during propagationwhich might have resulted in the changes of the microsatellite profile.
In conclusion, this study has established that the fungal inoculumused for seedling and flower inoculations has resulted in systemicendophytic infection similar to the endophytic infection of B. cinereapreviously reported in Primular polyantha [2]. As reported from theprevious work [21] which confirmed seed as the source of systemic B.cinerea infection. Therefore, resultsof the present study showed thatinfection in seedling grown from infected seed can be transferred intothe seed and subsequently to the seed of the next-generation plantwithout showing symptom a complete endophytic infection. This wasshown by re-isolation of the inoculum added to the plant from fewseeds isolates of first and second generation plant grown from nonsterilized seed at controlled environment room [22-24].
References
- Elias SNK, Shaw WM, Dewey FM (2010) Persistent symptomless, systemic, and seed-borne infection of lettuce by Botrytis cinerea. Eur J Plant Pathol 126: 61-71.
- Agrios GN (1997) Plant pathology: London press.
- Elad Y, Williamson B, Tudzynski P, Delen N (2004) Botrytis spp. and diseases they cause in agricultutal systems- An introduction. In Y. Elad (eds). Botrytis: Biology, Pathology and Control. 1-8 Springer, Dordrecht, The Netherland.
- Keller M, Viret O, Cole FM (2003) Botrytis cinerea infection in Grape flowers: Defence, Latency and Disease Expression. Phytopathology 9: 316-322.
- Koycu ND, Ozer N (1997) Determination of seedborne fungi in Onion andtheir transmission to Onion Sets. Phytoparasitica 25: 25-31.
- Barnes SE, Shaw M (2003) Infection of commercial hybrid primula seed by Botrytis cinerea and latent disease through the plants. Phytopathology 93: 573-578.
- Yourman LF, Jeffers SN, Dean RA (2000) Genetic analysis of isolates of B. cinerea sensitive and resistant to benzimidazole and dicarboximide fungicides. Phytopatology 90: 851-859.
- Burgess DR, Bretag TW, Keane PJ (1997) Seed to seedling transmission of Botrytis cinerea chickpea and disinfestation of seed with moist heat. Aust JExp Agric 37: 223-229.
- Denielson S, Mercado VH, Ames T, Munk S (2004) Seed transmission of downy (Perenosporra farinose f sp. Chenopodii) inqunoa and effect of relative humidity on seedling infection. Seed Science and Technology 32: 91-98.
- Rajaguru BAP (2008) Molecular ecology of Botrytis cinerea. PhD thesis. University of Reading, Reading, UK.
- Fournier E, Giraud A, Loiseau A, Vantrin D, Estoup A, et al. (2002) Characterization of nine polymorphic microsatellite Loci in the fungus Botrytis cinerea (Ascomycota). Molecular Biology Ecology 2.
- Williamson B, Tudzynski B, Tudzynski P, Van Kan JAK (2007) Botrytiscinerea: the cause of grey mould diseases. Mol Plant Pathol 8: 561-580.
- Singh D, Mathur SB (1992) Systemic infection of maize (Zea mays) seedlingsby Botryodiplodia theobromae. Indian Journal of Agricultural Sciences 62:713-716.
- Samir D, Amnon L (2007) Post-harvest Botrytis infection: Etiology, Development and Management. In Y. Elad, (eds) Botrytis: Biology Pathology and Control 349-367. Springer Dordrecht, The Neatherland.
- Koycu ND, Ozer N (1997) Determination of seedborne fungi in Onion and their transmission to Onion Sets. Phytoparasitica 25: 25-31.
- Ochoa JB, Ellis MA (2002) Seed transmission of Fusarium oxysporum in common naranjilla (Solanum quitoense) in Ecuador. Plant Health Progress.
- Shafia A (2009) Latent infection of Botrytis cinerea. PhD thesis, School of Biological Sciences. University of Reading.
- Marlatt RB, Lewis RW, McKittrick RT (2003) Systemic infection of Lettuce by Bremia lactucae. Phytopathology 52: 888-890.
- Suarz MB, Walsh K, Boonham N, O Neil T, Pearson S, et al. (2005) Development of real time PCR (Taqman) assays for the detection and quantification of B. Cinerea in planta. Plant Physiol Biochem 43: 890-899.
- Shah DA, Bergstrom GC (2002) Temperature dependent seed transmission of Stagnospora nodorum. Eur J Plant Pathol 106: 837-842.
- Sowley ENK (2006) Epidemiology of Botrytis cinerea in lettuce. PhD thesis , School Of Biological Sciences University of Reading.
- Viret O, Keller M, Jaudzems VG, Cole FM (2004) Botrytis cinerea infectionof Grape flowers: Light and electron microscopical Studies of infection sites.Phytopathology 94: 850-857.
- Holtz G, Williamson B (2004) The Ecology of Botrytis on plant Surfaces. InY. Elad (eds) Botrytis : Biology, Pathology and Control. Pp 9-27. Springer Netherland.
- Keller M, Viret O, Cole FM (2003) Botrytis cinerea infection in Grape flowers: Defence, Latency and Disease Expression. Phytopathology 9: 316-322.