Biotechnological Interventions and their Role in Sustainable Hill Agriculture
R. K. Salgotra*, B. B. Gupta and Monika Sood
Corresponding author: Dr. R. K. Salgotra, 1School of Biotechnology, Sher-e-Kashmir University of Agricultural Sciences &Technology of Jammu, Chatha, Jammu-190009 (J & K), India,; E-mail: rks_2959@rediffmail.com
1School of Biotechnology, Sher-e-Kashmir University of Agricultural Sciences & Technology of Jammu, Chatha, Jammu-190009(J & K), India
2Division of Plant Breeding & Genetics, Sher-e-Kashmir University of Agricultural Sciences & Technology of Jammu, Chatha,Jammu-190009 (J & K), India
3Division of Post Harvest Technology, Sher-e-Kashmir University of Agricultural Sciences & Technology of Jammu,Udheywala, Jammu-190018 (J & K), India
Citation: Salgotra RK, Gupta BB, Sood M. Biotechnological Interventions and their Role in Sustainable Hill Agriculture. J Plant Sci Res. 2015;2(1): 118.
Copyright © 2015 R. K. Salgotra. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Plant Science & Research | ISSN: 2349-2805 | Volume: 2, Issue: 1
Submission: 04/02/2015; Accepted: 13/03/2015; Published: 18/03/2015
Abstract
The ever increasing demand for food, feed, nutrition and other basic needs has put a lot of pressure on the agricultural and horticultural sectors in termsof addressing the accompanied challenges associated with increasing crop productivity in hills. At the same time, we are witnessing the potential benefitsof the revolutions forced by the science of biotechnology. Biotechnology has played an important role towards the attainment of production sustainabilityof crops by using various biotechnological tools. Biotechnology has the potential to provide new opportunities for achieving enhanced crop productivity ina way that will alleviate poverty, improve food security and nutrition, and promote sustainable use of natural resources in hill farming system. Conventionaland biotechnological research inputs have contributed in solving some of the constraints limiting crop productivity. However, limitations such as complexgenome, narrow genetic base, poor fertility, susceptibility to biotic and abiotic stresses and long duration to breed elite cultivars, hinder crop improvementprogrammes. Several radiation induced mutants with agronomically desirable traits were isolated and evaluated under field conditions, besides studyingabiotic stress responses using molecular tools. This article describes the developments in the in vitro culture systems and related biotechnologies tools suchas marker-assisted selection (MAS), marker-assisted breeding (MAB), QTL mapping etc. that are evolving as novel strategies in the recent years for use incrop improvement.
Keywords: Biotechnological strategies; Sustainability; Crop production; Hills
Introduction
The world’s population has increased from 2.5 billion to 6.4 billion in the last 50 years and the population explosion threatens that another 3 billion people will inhabit the planet by 2100. Each night 854 million people go to bed hungry and about 842 million people suffer from malnutrition [1]. It could be very difficult to improve nutritional deficiency exclusively by traditional breeding and management practices of crops. Efforts are being made towards nutritional improvement of crops by using the tools of biotechnology by increasing the levels of essential nutrients. A major challenge of sustainable livelihood in the developing nations can be met with the judicious and rational application of biotechnology along with other equally powerful crop production systems. Adoption of biotechnology in agriculture has been the focus of controversy due to varied reasons, as questions have arisen regarding food and environmental safety. The food safety assessment requires the evaluation of safety of newly added DNA and the safety of their gene products and overall safety of balance of food. The concept of substantial equivalence has been accepted as the cornerstone of the health hazard assessment of genetically modified (GM) foods. Therefore evaluation of such interventions necessitates establishing the reasonable likelihood of safety and that new varieties are as safe as or safer than the crops produced through traditional methods.
Agriculture in the hills is facing multifarious challenges in checking degradation of natural resources, developing adaptation and mitigation strategies in the changing climatic scenario and in meeting the changing food requirement for the increasing population. The constraints of contraction of the land holding size, local land tenure systems, undulating hilly topography further complicates the challenges making them more difficult to address. On the other hand, hills are endowed with rich repository of biological diversity, valuable genetic resources of agricultural crops and plethora of natural resources. The Green Revolution, launched in the 1960s and 1970s, used advances in science and technology to double and/or triple the yields of rice, wheat, and maize in Asia, saving hundreds of millions of lives. However, the Green Revolution bypassed hills. For hills to realize the benefits of a Green Revolution, a lot of effort, commitment and focus are required. Hills generally lacks the instruments of the Green Revolution development and use of high-yielding varieties, improved utilization of water through irrigation systems, utilization of farm inputs such as fertilizer and pesticides. To move forward, hill agriculture will not only need to address these aspects but also to recognize the new challenges the world is facing and new technological advancement presented by biotechnology.
To ensure food and nutritional security, sustainable development of hill agriculture through conservation and judicious utilization of natural resources, regional planning and demand driven technology led agriculture is essentially needed. The realistic utilization of biotechnology in agriculture has not only brought about profound changes in the crop productivity and quality, but has also opened up newer and hitherto unforeseen potential vistas including engineering novel traits, successful commercial micropropagation, manipulating plant architecture and molecular farming. The molecular techniques for plant diseases diagnosis and pathogen characterization, markerassisted selection, parasitic diagnosis, development of transgenic crop, and gender friendly agri-entrepreneurship have found a stay in the economic development and prosperity of farmers in the hilly region. In this chapter, we are reviewing the development challenges facing in hill agriculture and try to elucidate how biotechnology can contribute to food security, poverty reduction and sustainable socioeconomic development.
Biotechnological strategies to improve food security
Conventional and biotechnological research inputs have contributed in solving some of the constraints limiting crop productivity. However, limitations such as complex genome, narrow genetic base, poor fertility, susceptibility to biotic and abiotic stresses and long duration to breed elite cultivars, hinder crop improvement programmes. The following are the main biotechnological strategies that are being adopted to solve the biotic and abiotic stresses to enhance the crop productivity on sustainable basis.
Molecular marker technology
Isozyme and molecular based research accelerate the conventional plant breeding. DNA markers are the most widely used type of marker predominantly due to their abundance. They arise from different classes of DNA mutations such as substitution mutations (point mutations), rearrangements (insertions or deletions) or errors in replication of tandemly repeated DNA [2]. These markers are selectively neutral because they are usually located in non-coding regions of DNA. Unlike morphological and biochemical markers, DNA markers are practically unlimited in number and are not affected by environmental factors and/or the developmental stage of the plant [3]. Increasingly, techniques are being developed to more precisely, quickly and cheaply assess genetic variation. In recent years different molecular markers include: (i) hybridization-based markers such as restriction fragment length polymorphism (RFLP), (ii) PCRbased markers: random amplification of polymorphic DNA (RAPD), amplified fragment length polymorphism (AFLP) and microsatellite or simple sequence repeat (SSR), and (iii) sequence-based markers: single nucleotide polymorphism (SNP) have been developed andapplied to a large crop plant species. Now these molecular markers are being widely used to track loci and genome regions in several crop breeding programmes, as molecular markers tightly linked with a large number of agronomic and disease resistance traits are available in major crop species [4-6]. The majority of these molecular markers have been developed either from genomic DNA libraries (RFLPs and SSRs) or from random PCR amplification of genomic DNA (RAPDs) or both (AFLPs). These DNA markers can be generated in large numbers and can prove to be very useful for a variety of purposes relevant to crop improvement. Their association with genes/QTLs controlling the traits of economic importance has also been utilized in some cases for indirect MAS. Other uses of molecular markers include gene introgression through backcrossing, germplasm characterization, genetic diagnostics, characterization of transformants, study of genome organization and phylogenetic analysis [5].
Characterization, fingerprinting and identification
Different crop species e.g. rice, winter wheat, buckwheat, pigeon pea, beans, barley etc are characterized using isozymes. DNA molecular markers (RAPD, SSR) can be used to characterize and fingerprint the varieties and landraces of rice, barley, beans and buckwheat. Isozyme profiles of pigeon pea; RAPD profiles of winter wheat and buckwheat and SSR profiles of rice and barley has been done successfully. Isozyme and DNA profiles of different accessions can be developed. For identification, similarity or dissimilarity bands, private alleles or standard genotyping can be used. These markers can also be used to identify variety, F1 hybrid and in plant variety protection.
Screening and diversity study
It is planned to use DNA markers (RAPD, SSR) to screen the accessions for particular trait e.g. blast resistance and susceptible rice accessions, drought tolerance accessions, cold tolerance accessions, high protein content accessions etc. Diversity is assessed among variety and landraces of rice, barely, buckwheat, winter wheat, beans, etc using morphological, isozymes, RAPD and SSR markers. Different diversity parameters are estimated. Clustering is the common method of showing relationship based on the genetic distance. These findings are very useful for parent selection and heterosis prediction.
Linkage map construction and gene tagging
Linkage maps based on either morphological, isozymes or DNA markers are prerequisite to identify and tag desired genes. Mapping and tagging agriculturally important genes with molecular markersis the foundation for markers assisted selection (MAS) and marker assisted backcrossing (MAB) in crop plants. Molecular tags, a prerequisite for MAS and MAB have been developed for many crop plants using different kinds of molecular markers elsewhere. Linkage map on common buckwheat using EST markers on F2 population has been constructed. It is planned to construct such map in other important crop species. Tagging blast resistance gene in rice landraces of hills is expected and it will be used for MAS.
Marker assisted selection (MAS)
MAS provides new solutions for selecting and maintaining desirable genotype. Once molecular markers closely linked to desirable traits are identified, MAS can be performed in early segregating population and at early stages of plant development. Marker assisted selection or identification can be used to pyramid the major genes including resistance genes with the ultimate goal of producing varieties with more desirable characters. With MAS it is now possible for breeder to conduct many rounds of selection in a year. Molecular marker technology is now integrated into existing plant breeding programmes all over the world in order to allow researchers to access, transfer and combine genes at a rate and with a precision not previously possible. The potential for accelerating backcross breeding is possible now after the advent of DNA markers. These markers can be used to select backcross progeny which had usually large portions of recurrent parent genotype reducing the number of backcross generation necessary to reconstitute a recurrentparent genotype. MAB has been very widely used by commercial firm savings of even 1-2 generations.
QTL mapping and gene pyramiding
Most of the important agronomic characters like yield and yield components, plant height and days to flowering are controlled by several genes. A number of methods for mapping quantitative trait loci (QTL) and estimating their effects have been suggested and investigated. Many QTL can be identified using DNA markers. The identified QTLs can be used in MAS or MAB programmes in crop improvement of rice for various traits like drought tolerance accessions, cold tolerance accessions, high protein content accessions etc.
p> Potential application of biotechnology for sustainabledevelopment in hills
Agriculture is the mainstay of the economies in hill areas, to attain improved nutritional well-being and sustainable food security, emphasis should be put on strategies that will reduce the problems of poverty and underdevelopment because these are the underlying impediments to adequate access to food by all individuals. Crop improvement using various biotechnological tools is recognized globally as drivers of increased crop productivity and improving standards of living of farmers in hills. There is ample evidence that can solve the various problems of the farmers with the advances in agriculture biotechnology. Efforts are being made towards improvement of crops by using the tools of biotechnology on sustainable basis along with other crop production systems.
Application of tissue culture for propagation of virusfreeplanting material of vegetatively propagated andneglected/orphan crops of hills
Tissue culture is seen as a main technology for the large scale production of disease-free, high-quality planting material of crops such as potato, flowers and fruit crops. In commercial applications,such as fruit-export production and floriculture, it also generates much-needed employment, particularly for women. These techniques are very appropriate particularly for hills because they are relatively simple and inexpensive. Plant regeneration can occur through two main routes, direct and indirect morphogenesis. In direct morphogenesis, plants are regenerated directly from tissues such as immature leaf roll discs and also from shoot tip culture, by which it is propagated commercially. Indirect morphogenesis involves initial culturing of leaf roll sections or inflorescences on an auxin-containing medium to produce an undifferentiated mass of cells, or callus. Somaticembryogenesis techniques have two main goals: the development of a highly efficient method for propagating large number of uniform plants in less time and possibly at lesser cost than the conventional propagation methods, and a cell culture based regeneration system useful for genetic transformation [7]. Embryogenic cultures have also found their place in a wide variety of applications, from obtaining virus resistant plants through somaclonal variation, to mutagenesis and in vitro selection and developing transgenic plants.
Induced mutations have contributed significantly and several mutant varieties have been developed through mutation breeding. In vitro culture combined with radiation induced mutagenesis in the improvement of crops. Somaclonal variation in combination with in vitro mutagenesis can be beneficial for the isolation of salinity and drought tolerant lines in a short duration employing in vitro selection. Earlier studies using radiation induced mutagenesis and in vitro techniques led to the development of varieties using 5, 20, 40 and 60 Gy gamma-ray irradiated cultures [8].
Use of marker technology for important food crops
With the advent of molecular marker technology, several types of molecular markers were developed to assist in selection of desired trait in the segregating and advanced breeding generations. In current scenario, the DNA markers become the marker of choice for the study of crop genetic diversity has become routine, to revolutionized the plant biotechnology. DNA markers could be used to identify traits in plants such as maize, rice, wheat, apple, pears, plums etc. widely grown in hills. These markers assist in the development of new breeds/crop varieties, which could be resistant to biotic and abiotic stresses. Marker technology is appropriate because genes associated with important traits can now be detected by simple laboratory tests in embryos and/or young plantlets. It is therefore no longer necessary to grow plants to maturity in the field to find out whether or not they possess a particular trait. As such, the time it takes to develop a new variety could be greatly reduced through the use of accurate markers for a gene or genes. Molecular markers are also used to analyze genetic variability or to assess the relationships between populations or gene pools. This helps in the search for potentially useful materials, which could be used in breeding, or to decide on conservation measures for threatened crop and neglected/orphan crops.
With the advent of molecular markers in the late 1970s markerassisted selection (MAS), a breeding approach based on the genotype of plants rather than assessing the phenotype only, became possible. From the beginning, the expectations about the potentials of MAS have been huge. Nevertheless, there is little systematic knowledge about the degree to which MAS is being used in modern plant breeding programs and for what purposes. MAS offered possibilities to plant breeders to overcome many of the problems faced during conventional breeding. Knowledge of the location of all genes in a genome extends molecular marker technology because it becomes possible to identify candidate genes controlling specific traits. The genes then become the markers and the process becomes more accurate and more efficient. The location and sequence of candidate genes makes it possible to design allele specific markers which readily lend themselves to automation. In biotechnology, markerassisted selection (MAS) and marker-assisted breeding (MAB) has the potential to increase crop productivity in hill farming system innumber of crops (Table 1).
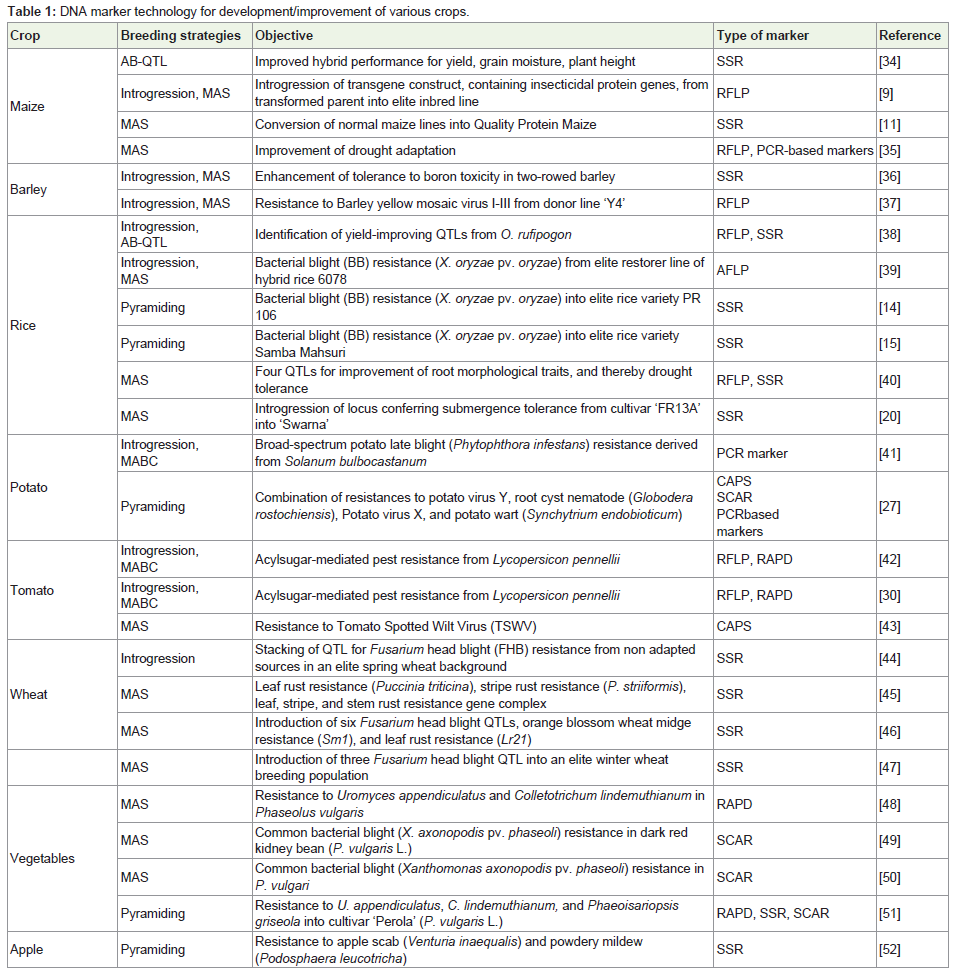
Table 1: DNA marker technology for development/improvement of various crops.
Maize
Maize (Zea mays L.) is an important food and feed crop of the world. It ranks fifth in acreage and third in production. It is one of the major sources of calorie and protein. However, it is deficient in essential amino acids viz., lysine and tryptophan. Quality protein maize(QPM) with opaque-2 gene along with associated modifiers contains twice as much lysine and tryptophan and 30 per cent less leucine than the normal maize. The reduced level of zein further improves the nutritional quality of the QPM. Among the cereals, maize is the first crop to which MAS is being applied on a large scale. Maize varieties are predominantly F1 hybrids and breeding in the developed countries is dominated by a small number of large private sector breeding companies [9]. According to Ragot and Lee [10] the most important use of MAS in maize is backcrossing of transgenes into elite inbred lines. Other important examples for the successful application of MAS in maize are the use of microsatellite markers for the conversion of normal maize lines into Quality Protein Maize (QPM), containing more lysine and tryptophan than the native lines [11], or the introgression of favorable QTL for earliness and grain yield between maize elite lines [12].
Rice
Rice, having the smallest genome of all cultivated cereals, being diploid and self pollinating, is the most extensively studied species among cereals. Up to now, MAS in rice breeding has mainly been utilized for the pyramiding of disease resistances, namely bacterial blight [13-15] and blast [16]. In 2002, two cultivars resistant to bacterial leaf blight were released in Indonesia, which have been selected using MAS. The variety ‘Angke’ carries the resistance gene xa5, ‘Conde’ carries Xa7 [17]. Several publications report introgressions from wild relatives (e.g. O. glumeapatula, O. rufipogon) in order to improve yield [18,19] and in 2006, two lines showing strong submergence tolerance were developed by introgressing a locus conferring submergence tolerance from cultivar ‘FR13A’ into the variety ‘Swarma’ [20]. In India, two rice varieties such as Improved Pusa Basmati 1 and Improved Samba Mahsuri has been developed through MAS and are being widely grown.
Barley
Barley is, like wheat, a self-pollinating species and in terms of the breeding system and the economic structure of its market it resembles wheat. However, MAS seems to have progressed further than in wheat, which is probably due to the simpler, diploid genome. A main focus of marker selection in barley is breeding for barley yellow mosaic virus resistance; another important focus is rust resistance. In contrast to wheat, barley varieties have been released that are based on MAS. In the USA the variety ‘Tango’, carrying two QTL for adult resistance to stripe rust, was released in 2000 [21], claiming to be the first commercially released barley variety using MAS. However, ‘Tango’ yields less than its recurrent parent and is therefore primarily seen as a genetically characterized source of resistance to barley stripe rust rather than a variety of its own. As a result of the South Australian Barley Improvement Program the malting variety ‘Sloop’ was improved with cereal cyst nematode resistance introgressed from the variety ‘Chebec’ and released in 2002 as ‘Sloop SA’ [22].
Legumes
Legumes are the major food crops grown and consumed mostly by the poor farmers as an important source of protein in the drylands of the world. Despite many uncertainties, there is a growing consensus that the climatic adversities could lead to an overall increase in the disease and pest pressure. Since most of the legumes have a narrow genetic base and levels of resistance to some biotic and abiotic constraints are low, making crop improvement an overarching research for development challenge for maximizing the benefits to small farmers in hills. Running against the headwinds, grain legume research has been immensely benefited by applications of modern biotechnological tools and approaches that have the potential to develop solutions for destructive diseases, besides making headway against the complex problems of drought [53]. Similarly, identifying novel genes/traits and assessing their suitability as candidate genes for genetic engineering options will be important for future breeding programs in order to achieve remarkable impacts in these grain legume crops globally. This chapter mainly provides a comprehensive picture of the different biotechnological interventions adopted for addressing various constraints in gain legume productivity and improvement, highlighting the pitfalls and possible solutions that can be taken through an integrated approach to combat the altered environmental conditions.
Potato
Potato (Solanum tuberosum L.) is the most important non-cereal food item worldwide. It produces more dry matter and calories per unit land and time than any other major food crop. Potatoes are the world’s most important vegetable and fourth most important foodcrop in terms of production [23]. Potato is one of the first major crops where biotechnology has been successfully employed. With the application of biotechnology, disease resistant, insect pest resistant, and herbicide tolerant plants have been produced. These advancesare only the beginning of a second green revolution and coupled with conventional breeding will lead to ‘low input’ agriculture by proving farmers with ‘tailored’ seeds which can protect themselves against biotic and abiotic stress. This would also reduce the amount of hazardous chemicals in the environment and foodstuffs. Nevertheless, MAS has been practiced only little because the construction of classical genetic linkage maps was not practical for a long time, due to the tetraploidy of potatoes combined with tetrasomic inheritance. This complicates the detection of linkages and largely prevents the recovery of recessive phenotypes. Only with the technical developments that allowed to reduce ploidy from the tetraploid to the diploid level and with the advent of DNA-based markers it became possible to construct detailed genetic maps for potato and to carry out genome-wide studies on qualitative and quantitative disease resistance [24].
Several linkage maps have been constructed with molecular markers over the past 20 years and the most important marker applications in potato breeding are marker assisted resistance breeding and marker-assisted introgression [25]. Although various markers have been developed [25], practical applications in breeding are still rare. Most marker applications are carried out in experimental populations of diploid potatoes [26], which impedes the application of experimental results in practical breeding work [27]. The authors applied molecular markers to develop breeding material that carries resistances to three different pathogens and the root cyst nematode (Globodera rostochiensis).
Tomato
Tomato is one of the most important vegetable crop worldwide and according to Foolad and Sharma [28] and MAS has mainly been utilized in resistance breeding. However, MAS is not yet a routine procedure for improving quantitative traits, although it has been employed to improve quantitative resistance to bacterial wilt, bacterial cancer and tomato yellow leaf curl virus as well as for improving fruit soluble solids.†In tomato breeding programs, the useof MAS is less common, although MAS has been used to improve horizontal resistances to black mold [29] and for some complex fruitqualitycharacteristics [30].
Beans and other vegetables
Marker application in vegetable breeding programs comprises lettuce, pepper, cucumber, and several Phaseolus spp.of beans. Total of 25 alleles in nine loci formed two groups showing intra and inter populations variations in diversity study at molecular level of Phaseolus spp. And landraces showed more variations than improved. RAPD and SCAR markers were used predominantly for MAS for disease resistance in vegetables [31]. MA was also used for improvement of varieties for disease resistances like resistant to bean golden yellow mosaic virus [32].
Fruits
Many fruit crops like the pome fruits have a large size, long juvenile phase, and a high level of heterozygosity which makes classical genetic analysis a difficult task. The cost-benefit relationships of MAS are more favorable for fruit crops than for annual crops because most fruit breeding programs consist of two stages. In the first stage, large populations of non replicated individuals are evaluated and only a small number of plants are selected for extensive asexual propagation to be tested in replicated trials in the second breeding stage. Especially during the first stage the large plant size and long life cycle have a negative impact. With the aid of MAS numbers of plants that have to be tested in the first breeding stage can be drastically reduced.
However, many important agronomic characters in fruit trees are related to major genes, e.g. several disease resistances, flower characteristics, and fruit or nut quality traits [33] and very few QTLs are available in fruit breeding. MAS can be applied successfully in fruit breeding now and more applications are to be expected, bearing in mind the tremendous efficiency gain which molecular markers provide in fruit breeding.
Biotechnology for bio-processing and value addition
A significant amount of the food produced is lost after harvest thereby aggravating hunger. Most crops such as apple, pear and potatoes are perishable crops with a lot of storage and post-harvestproblems. Productivity is limited by their rapid rate of deterioration soon after harvesting, if processing into more shelf-stable products is delayed. Estimates of post-harvest losses are hard to determine, but some authorities put losses to as high as 50 percent of what is produced. It is therefore, important to develop and deploy crop varieties with improved shelf life.
Biotechnology can also be used to convert agricultural waste into useful products such as industrial fibres, bioethanol, wine, beers, animal feeds, fertilizers, cosmetics and pharmaceuticals. We should use biotechnology to generate new technologies and up-scale known and proven technologies for value addition, reducing and/or eliminating post-harvest losses in the hills. Technologies for processing and/or value addition are available but not yet commercially transferred to even progressive farmers.
Genetic engineering
Like marker-assisted breeding, GM technology has the potential to increase crop productivity and food security more efficiently through the development of improved varieties with increased yields, nutritional content and storage characteristics. Countries like India should continue to build capacity in GM technology, giving priority to traits and crops that are of economic importance such as drought tolerance in food crops such as maize, rice and sorghum, salinity or flood tolerance in rice, increased nutrition value and improved shelf life. Stronger and more diversified collaborations between and among the public sector, private industry and civil society - coupled with the appropriate policy environment and organizational structures are needed to influence the formation of effective innovation systems that will turn ideas into products and services. Currently India is growing only Bt cotton crop at commercial scale. Although GM brinjal is also in pipelines but its fate of commercial use is still debatable. In India R & D projects are going on for development of GM in number of crops species. In recognition of the role of GM technology in contributing to food security and poverty reduction a number of countries have established R&D programmes in genetic engineering (Table 2).
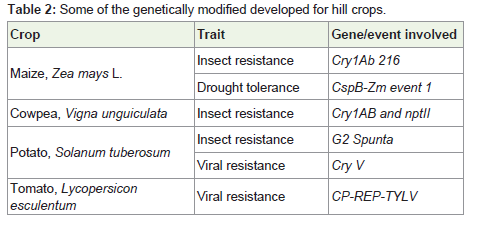
Table 2: Some of the genetically modified developed for hill crops.
Conclusion
Biotechnology is having great opportunities for the increase in agricultural production globally. Biotechnology has played an important role towards the attainment of production sustainability of crops by using various biotechnological tools. Increasing global food production within existing land area and the use of modern plant breeding methods have enhanced increased production of crops like legumes to improve soil structure, organic matter and fertility. These lead to conservation of bioresources and prevent soil erosion in hills. Agricultural biotechnology, including tissue culture, DNAmarker- assisted breeding and GM technology, has the potential to increase crop productivity and food security in hills more efficiently through the development of improved varieties with increased yields, nutritional content and storage characteristics. Biotechnological tools should continue to build capacity in GM technology, giving priority to traits and crops that are of economic importance such as drought tolerance in food crops such as maize, rice, beans, cold tolerance in rice, increased nutrition value and improved shelf life. Stronger and more diversified collaborations between and among the public sector and private industry are needed to influence the formation of effective innovation systems that will turn ideas into products for the benefit of farmer community.
References
- Food and Agriculture Organization (2006) The State of Food and Agriculture.
- Paterson AH (1996) Making genetic maps. In: A.H. Paterson (Ed.), Genome Mapping in Plants, pp. 23-39. R.G. Landes Company, San Diego, California; Academic Press, Austin, Texas.
- Winter P, Kahl G (1995) Molecular marker technologies for plant improvement. World J Microbiol Biotechnol 11: 438-448.
- Phillips R L and Vasil I K (2001) DNA-based markers in plants. In: Phillips RL, Vasil IK (eds) DNA-based markers in plants. Kluwer Academic Publishers, Dordrecht, The Netherlands, p 497.
- Jain SM, Brar DS, Ahloowalia BS (2002) Molecular techniques in crop improvement. Kluwer Academic Publishers, The Netherlands.
- Gupta PK, Varshney RK (2004) Cereal genomics: an overview In: Gupta PK, Varshney RK (eds) Cereal genomics. Kluwer Academic Press, Dordrecht, The Netherlands, p 639.
- Lakshmanan P, Geijskes RJ, Aitken KS, Christopher LP, Grof Bonnett GD, et al. (2005) Sugarcane Biotechnology: the challenges and opportunities. In Vitro Cell Dev Biol Plant 41: 345-363.
- Patade VY, Suprasanna P, Bapat VA (2008) Gamma irradiation of embryogenic callus cultures and in vitro selection for salt tolerance in sugarcane (Saccharum offcinarum L.) Agril Sci in China 7: 1147-1152.
- Ragot M, Biasiolli M, Delbut M F, Dellorco A, Malgarini L, Thehevnin P, Vernoy J, Vivant J, Zimmermann R and Gay G (1995) Marker-assisted backcrossing: A Practical Example. - In: Bervillé, A. & Tersac, M. (Eds.): Les Colloques, No. 72, Techniques et utilisations des marqueurs moléculaires. Montpellier (INRA) pp. 45-56.
- Ragot M and Lee M (2007) Marker-Assisted Selection in Maize: Current Status, Potential, Limitations and Perspectives From the Private and Public Sectors. - In: Guimaraes, E. P., Ruane, J., Scherf, B. D., Sonnino, A. & Dargie, J. D. (Eds.): Marker-assisted Selection - Current Status and Future Perspectives in Crops, Livestock, Forestry and Fish. Rome (Food and Agriculture Organization of the United Nations pp. 117-150.
- Babu ER, Mani VP, Gupta HS (2004) Combining high protein quality and hard endosperm traits through phenotypic and marker assisted selection in maize. Proceedings of the 4th International Crop Science Congress, Brisbane.
- Bouchez A, Hospital F, Causse M, Gallais A, Charcosset A (2002) Marker-assisted introgression of favorable alleles at quantitative trait loci between maize elite lines. Genetics 162: 1945-1959.
- Chen S, Lin X H, Xu C G and Zhang Q (2000) Improvement of bacterial blight resistance of `Minghui 63', an elite restorer line of hybrid rice, by molecular marker-assisted selection. Crop Sci 40: 239-244.
- Singh S, Sidhu JS, Huang N, Vikal Y, Li Z, et al. (2001) Pyramiding three bacterial blight resistance genes (xa5, xa13 and Xa21) using marker-assisted selection into indica rice cultivar PR106. Theor Appl Genet 102: 1011-1015.
- Sundaram RM, Sakthivel K, Hariprasad AS, Ramesha MS, Viraktamath BC, et al. (2010) Development and validation of a PCR-based functional marker system for the major wide-compatible gene locus S5 in rice. Mol Breed 26: 719-727.
- Narayana NN, Baishkh N, Oliva NP, Veracruz CM, Gnanamamickam SS, et al. (2004) Molecular breeding: Marker-assisted selection combined with biolistic transformation for blast and bacterial blight resistance in indica rice (cv. CO39). Mol Breed 14: 61-71.
- Bustamam M, Tabien R E, Suwarno A, Abalos M C, Kadir T S, Ona I, Bernardo M, Veracruz C M and Leung H (2002) Asian Rice Biotechnology Network: Improving Popular Cultivars Through Marker-Assisted Backcrossing by the NARES. - Poster presented at the International Rice Congress, 2002 September 16-20, Beijing, China.
- Brondani C, Rangel N, Brondani V, Ferreira E (2002) QTL mapping and introgression of yield-related traits from Oryza glumaepatula to cultivated rice (Oryza sativa) using microsatellite markers. Theor Appl Genet 104: 1192-1203.
- Liang F, Deng Q, Wang Y, Xiong Y, Jin D, et al. (2004) Molecular marker-assisted selection for yield enhancing genes in the progeny of "9311× O. rufipogon" using SSR. Euphytica 139: 159-165.
- Xu K, Xu X, Fukao T, Canlas P, Maghirang-R0driguez R, et al. (2006) Sub1A is an ethylene response factor like gene that confers submergence tolerance to rice. Nature 442: 705-708.
- Hayes PM, Corey AE, Mundt C, Toojinda T, Vivar H (2003) Registration of 'Tango' Barley. Crop Sci 43: 729-731.
- Eglinton JK, Coventry SJ, Chalmers K (2006) Breeding outcomes from molecular genetics. Breeding for success: Diversity in action: Proceedings of the 13th Australasian Plant Breeding Conference, 18-21 April, 2006, Christchurch, New Zealand: 743-749.
- Simko I (2004) One potato, two potato: Haplotype association mapping in autotetraploids. Trends Plant Sci 9: 441-448.
- Gebhardt C (2004) Potato genetics: Molecular maps and more. - In: Lorz, H. and Wenzel, G. (Eds.): Biotechnology in Agriculture and Forestry, Vol. 55, Molecular Marker Systems. - Berlin (Springer-Verlag): 215-227.
- Vreugdenhil D, Bradshaw J, Gebhardt C, Govers F, Mackrron D (2007) Potato biology and biotechnology: Advances and Perspectives. - Amsterdam (Elsevier Science & Technology).
- Gebhardt C (2007) Molecular markers, maps and population genetics. In: Lorz, H. and Wenzel, G. (Eds.): Biotechnology in Agriculture and Forestry, Vol. 55, Molecular Marker Systems. Amsterdam (Elsevier): 77-89.
- Gebhardt C, Bellin D, Henselewski H, Lehmann W, Schwarzfischer J, et al. (2006) Marker-assisted combination of major genes for pathogen resistance in potato. Theor Appl Genet 112: 1458-1464.
- Foolad MR, Sharma A (2005) Molecular markers as selection tools in tomato breeding. Acta Horticulturae 695: 225-240.
- Robert VJM, West MAL, Inai S, Caines A, Arntzen L, et al. (2001) Marker-assisted introgression of blackmold resistance QTL alleles from wild Lycopersicon cheesmanii to cultivated tomato (L. esculentum) and evaluation of QTL phenotypic effects. Mol Breed 8: 217-233.
- Lecomte L, Duffe P, Buret M, Servin B, Hospital F, et al. (2004) Marker-assisted introgression of five QTLs controlling fruit quality traits into three tomato lines revealed interactions between QTLs and genetic backgrounds. Theor Appl Genet 109: 658-668.
- Fan Z, Robbins M, Staub J (2006) Population development by phenotypic selection with subsequent marker-assisted selection for line extraction in cucumber (Cucumis sativus L.). Theor Appl Genet 112: 843-855.
- Beaver J S, Porch T G and Zapata M (2008) Registration of 'Verano' White Bean. J Plant Registrations 2: 187-189.
- Dirilewanger E, Graziano E, Joobeur T, Garriga-Caldere F, Cosson P, et al. (2004) Comparative mapping and marker-assisted selection in rosaceae fruit crops. Proceedings of the National Academy of Sciences of the United States of America 101: 9891-9896.
- Ho C, Mccouch R, Smith E (2002) Improvement of hybrid yield by advanced backcross QTL analysis in elite maize. Theor Appl Genet 105: 440-448.
- Ribaut J M, Ragot M (2006) Marker-assisted selection to improve drought adaptation in maize: The backcross approach, perspectives, limitations, and alternatives. J Exp Bot 58: 351-360.
- Emebiri L, Michael P, Moody D (2009) Enhanced tolerance to boron toxicity in two-rowed barley by marker-assisted introgression of favourable alleles derived from Sahara 3771. Plant and Soil 314: 77-85.
- Okada Y, Kanatani R, Arai S, Asakura T, Ito K (2003) Production of a novel virus-resistant barley line introgression to the rym1 locus with high malting quality using DNA marker assisted selection. J of the Institute of Brew 109: 99-102.
- Xiao J, Li J, Grandillo S, Ahn SN, Yuan L, et al. (1998) Identification of trait-improving quantitative trait loci alleles from a wild rice relative, Oryza rufipogon. Genet 150: 899-909.
- Chen S, Xu CG, Lin XH, Zhang Q (2001) Improving bacterial blight resistance of' 6078', an elite restorer line of hybrid rice, by molecular marker-assisted selection. Plant Breed 120: 133-137.
- Steele KA, Price A, Shashidhar HH, Witcombe JR (2006) Marker-assisted selection to introgress rice QTLs controlling root traits into an Indian upland rice variety. Theor Appl Genet 112: 208-221.
- Colton LM, Groza HI, Wielgus SM, Jiang J (2006) Marker-assisted selection for the broad-spectrum potato late blight resistance conferred by gene RB derived from a wild potato species. Crop Sci 46: 589-594.
- Lawson DM, Lunde CF, Mutschler MA (1997) Marker-assisted transfer of acylsugar-mediated pest resistance from the wild tomato, Lycopersicon pennellii, to the cultivated tomato, Lycopersicon esculentum. Mol Breed 3: 307-317.
- Langella R, Ercolano M, Monti L, Frusciante L and Barone A (2004) Molecular marker assisted transfer of resistance to TSWV in tomato elite lines. J of Hort Sci Biot 79: 806-810.
- Miedaner T, Wilde F, Steiner B, Buerstmayr H, Korzun V, et al. (2006) Stacking quantitative trait loci (QTL) for Fusarium head blight resistance from non-adapted sources in an european elite spring wheat background and assessing their effects on deoxynivalenol (DON) content and disease severity. Theor Appl Genet 112: 562-569.
- Chicaiza O, Khan IA, Zhang X, Brevis JC, Jackson L, et al. (2006) Registration of five wheat isogenic lines for leaf rust and stripe rust resistance genes. Crop Sci 46: 485-487.
- Hodges DM, DeLong JM, Forney CF, Prange RK (1999) Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta. 207: 604-611.
- Somers D, Thomas J, Depauw R, Fox S, Humphreys G, et al. (2005) Assembling complex genotypes to resist Fusarium in wheat (Triticum aestivum L.). Theor Appl Genet 111: 1623-1631.
- Wilde F, Schon C, Korzun V, Ebmeyer E, Schmolke M, et al. (2008) Marker-based introduction of three quantitative-trait loci conferring Resistance to Fusarium head blight into an independent elite winter wheat breeding population. Theor Appl Genet 117: 29-35.
- Faleiro FG, Ragagnin VA, Moreira MA, De Barros E G (2004) Use of molecular markers to accelerate the breeding of common bean lines resistant to rust and anthracnose. Euphytica 138: 213-218.
- Miklas PN, Smith JR, Singh SP (2006a) Registration of common bacterial blight resistant white kidney bean germplasm line USDK-CBB-15. Crop Sci 46: 1005-1007.
- Miklas P N, Smith J R and Singh S P (2006b) Registration of common bacterial blight resistant white kidney bean germplasm line USWK-CBB-17. Crop Sci 46: 2338-2339.
- Raganin VA, Sanglard DA, De Souza TLPO, Moreira MA, De Barros EG (2003) Simultaneous transfer of resistance genes for rust, anthracnose and angular leaf spot to cultivar Perola assisted by molecular markers. Annual report of the bean improvement cooperative 46: 159-160.
- Kellerhals M, Szekely T, Sauer C, Frey J, Patocchi A (2009) Pyramiding scab resistances in apple breeding. Erwerbs-Obstbau 51: 21-28.
- Bhatnagar-Mathur, P, Palit P, Sridhar Kumar Ch, Reddy DS, Sharma KK (2012) Grain Legumes: Biotechnological Interventions in Crop Improvement for Adverse Environments. In: Improving Crop Productivity in Sustainable Agriculture. John Wiley & Sons, Inc: 381-421.