Research Article
Role of Cell Wall–Degrading Enzymes of Paradendryphiella arenariae, a Novel Pathogen in Fruit Rot Disease of Pomegranate
Krishnamurthi AV and Karigar CS
Department of Biochemistry, Bangalore University, Bangalore, Karnataka, India
*Corresponding author:Chandrakant S. Karigar, Department of Biochemistry, Bangalore University, Bangalore, Karnataka, India. E-Mail Id: karigar@bub.ernet.in
Copyright: © Krishnamurthi AV, et al. 2026. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Article Information:Submission: 13/05/2026; Accepted: 28/05/2026; Published: 30/05/2026
Abstract
Pomegranate (Punica granatum L.) is highly susceptible to fungal fruit rot, resulting in substantial yield and postharvest losses worldwide. Disease progression is largely mediated by extracellular cell wall–degrading enzymes (CWDEs), which facilitate host tissue maceration and pathogen invasion. In the present study, Paradendryphiella arenariae, isolated from naturally infected pomegranate fruits, was investigated for its CWDE production potential and represents the first report of this fungus associated with pomegranate fruit rot. Enzyme production was carried out under submerged fermentation using a pomegranate peel–based medium to simulate host-derived substrates. Qualitative plate assays confirmed the secretion of cellulase, xylanase, pectinase, lipase, laccase and proteases. Quantitative analysis revealed peak enzyme activities at day 14, with cellulase (11.8 U/mL), β-glucosidase (14.6 U/mL),
xylanase (10.0 U/mL), and pectinase (9.6 U/mL). SDS–PAGE profiling demonstrated multiple extracellular proteins in the range of 17–75 kDa, indicating a complex enzyme system. The predominance of pectinolytic, cellulolytic, and hemicellulolytic activities suggests a coordinated mechanism for degradation of fruit cell wall polysaccharides. These findings provide mechanistic insight into CWDE-mediated tissue disintegration during infection and a basis for
developing targeted management strategies to mitigate fungal fruit rot and reduce pre- and postharvest losses in pomegranate.
Keywords:Cell Wall Degrading Enzymes; Paradendryphiella arenariae; Cellulase; Β-Glucosidase; Xylanase; Pectinase; Hemicellulose; Laccase
Introduction
Fruit rot diseases caused by fungal pathogens represent a major
constraint on pomegranate (Punica granatum L.) production,
resulting in significant pre- and post-harvest losses that diminish
market value. These infections are caused by a host of fungal species
belonging to Alternaria, Aspergillus, Beltaraniella, Cercospora,
Cladosporium, Colletotrichum, Curvularia, Fusarium, Phomopsis,
Phytophthora, Rhizopus, etc. The infection process in fruit rot
pathogens largely depends on their ability to breach host structural
barriers, particularly the plant cell wall, which serves as the first line
of defence against microbial invasion. Successful colonization of fruit
tissues is strongly associated with the secretion of fungal cell walldegrading
enzymes (CWDEs), which facilitate tissue maceration [1,2], nutrient acquisition, and pathogen spread within the host.
The pomegranate fruit pericarp and aril tissues are rich in
complex polysaccharides such as cellulose, hemicellulose, and pectin,
along with phenolic compounds that contribute to host resistance.
The fruit-rot-causing fungi secrete a coordinated set of hydrolytic
enzymes, including cellulases, β-glucosidase, xylanases, pectinases
and proteases, which degrade the structural polysaccharides of the
host cell wall and overcome the barriers. Among these, pectinases play
a particularly critical role in fruit rot development by depolymerizing
pectin in the middle lamella, leading to cell separation, tissue softening
[3,4] and rapid fruit decay.
In addition to polysaccharide-degrading enzymes, oxidative
enzymes such as laccases contribute to the pathogenic process
by modifying lignin-like components, detoxifying host-derived
phenolic compounds [5,6] and facilitating fungal survival in hostile
host environments. The combined action of hydrolytic and oxidative
CWDEs enhances the virulence of fruit rot pathogens and determines
the severity and progression of disease symptoms.
Despite extensive reports on fungal fruit rot pathogens affecting pomegranate, information regarding the enzymatic mechanism employed by P arenariae during host infection is scarce. This dematiaceous fungus has been previously reported from marine and terrestrial environments, with limited evidence of its pathogenicity in economically important fruit crops. In the present study, P. arenariae is reported for the first time as a causal agent of natural pomegranate fruit rot. Understanding the enzymatic profile of P. arenariae and its role in pomegranate fruit rot provides valuable insights into the mechanisms of host-pathogen interaction and disease development. Such information is essential for elucidating the virulence attributes of newly emerging fruit rot pathogens and may contribute to the development of effective disease management and postharvest control strategies. Hence, in this report we characterize the key CWDEs and their role in pathogenesis in fruit rot disease.
Despite extensive reports on fungal fruit rot pathogens affecting pomegranate, information regarding the enzymatic mechanism employed by P arenariae during host infection is scarce. This dematiaceous fungus has been previously reported from marine and terrestrial environments, with limited evidence of its pathogenicity in economically important fruit crops. In the present study, P. arenariae is reported for the first time as a causal agent of natural pomegranate fruit rot. Understanding the enzymatic profile of P. arenariae and its role in pomegranate fruit rot provides valuable insights into the mechanisms of host-pathogen interaction and disease development. Such information is essential for elucidating the virulence attributes of newly emerging fruit rot pathogens and may contribute to the development of effective disease management and postharvest control strategies. Hence, in this report we characterize the key CWDEs and their role in pathogenesis in fruit rot disease.
Materials and Methods
Fungal isolate:
The fungal isolate Paradendryphiella arenariae used in the present
study was previously isolated from naturally infected pomegranate
fruits collected from Srinivaspura Taluk, Karnataka, and identified
based on detailed morphological and molecular characterisation in
our earlier report [7]. The culture was maintained on Potato Dextrose
Agar (PDA) slants at 4 °C and subcultured periodically to ensure
viability. The previously characterised isolate was used for subsequent
submerged fermentation, qualitative and quantitative enzyme
production studies.Qualitative Enzyme profile:
Cellulase: Cellulase activity was screened using carboxymethyl
cellulose (CMC) agar following the Cango red plate assay method
[8,9], The medium contained (g L-¹): NaNO₃, 2.0; KH₂PO₄, 1.0;
KCl, 0.5; MgSO₄·7H₂O, 0.5; FeSO₄·7H₂O, 0.01; agar, 15.0; and
carboxymethyl cellulose (CMC), 10.0 as the sole carbon source. Plates
were inoculated with the fungal culture and incubated at 28 °C for 7
days. After incubation, plates were flooded with 0.1% Congo red for
15 min and destained with 1 M NaCl for 10 min. Formation of clear
hydrolysis zones around the colony indicated cellulase activity. The
cellulolytic index (CI) was calculated as the ratio of hydrolysis zone
diameter to colony diameter.Xylanase: Xylanase activity was determined using xylan agar media [10]. The medium contained (g L-¹): NaCl (0.5), KH₂PO₄ (1.0), NH₄NO₃ (0.3), MgSO₄·7H₂O (0.5), FeSO₄·7H₂O (0.01), MnSO₄·H₂O (0.01), agar (20.0), and beechwood xylan (10.0) as the sole carbon source. Plates were inoculated with fungal cultures and incubated at 28 °C for 7 days. After incubation, plates were flooded with 0.1% Congo red solution for 15 min and destained with 1 M NaCl for 10– 15 min. The appearance of clear hydrolysis zones surrounding fungal colonies indicated xylanase activity.
Pectinase: Pectinase activity was qualitatively screened on pectin
agar [11,12] media. The medium contained (g L-¹): NaNO₃ (1.0), KCl
(1.0), K₂HPO₄ (1.0), MgSO₄·7H₂O (0.5), yeast extract (0.5), citrus
pectin (10.0), and agar (20.0), with the pH adjusted to 7.0 before
sterilisation. Plates were inoculated with 5 mm diameter fungal
mycelial discs and incubated at 28 °C for 7 days. After incubation,
plates were flooded with 0.1% Gram’s iodine solution for 5 min and
rinsed gently with distilled water. The formation of clear hydrolysis
zones surrounding the fungal colonies indicated pectinolytic activity.
Lipase: Lipase activity was qualitatively screened using tributyrin agar media [13]. The medium contained (g L-¹): peptone (5.0), yeast extract (3.0), NaCl (5.0), agar (15.0), and tributyrin (10 mL L-¹; 1% v/v). The pH of the medium was adjusted to 7.0 before sterilisation. Plates were centrally inoculated with 5 mm diameter fungal mycelial discs and incubated at 28 °C for 5-7 days. The formation of clear hydrolysis zones surrounding fungal colonies was recorded as a positive indication of extracellular lipase activity.
Amylase: Amylolytic activity was qualitatively screened on starch agar media [14]. The medium contained (g L-¹): peptone (5.0), yeast extract (3.0), NaCl (5.0), soluble starch (10.0), and agar (15.0). The pH of the medium was adjusted to 7.0 before sterilisation. Plates were centrally inoculated with 5 mm diameter fungal mycelial discs and incubated at 28 °C for 7 days. After incubation, the plates were flooded with Gram’s iodine solution for 5 min and rinsed gently with distilled water. The formation of clear hydrolysis zones surrounding the fungal colonies against a dark blue background was recorded as a positive indication of extracellular amylase activity.
Lipase: Lipase activity was qualitatively screened using tributyrin agar media [13]. The medium contained (g L-¹): peptone (5.0), yeast extract (3.0), NaCl (5.0), agar (15.0), and tributyrin (10 mL L-¹; 1% v/v). The pH of the medium was adjusted to 7.0 before sterilisation. Plates were centrally inoculated with 5 mm diameter fungal mycelial discs and incubated at 28 °C for 5-7 days. The formation of clear hydrolysis zones surrounding fungal colonies was recorded as a positive indication of extracellular lipase activity.
Amylase: Amylolytic activity was qualitatively screened on starch agar media [14]. The medium contained (g L-¹): peptone (5.0), yeast extract (3.0), NaCl (5.0), soluble starch (10.0), and agar (15.0). The pH of the medium was adjusted to 7.0 before sterilisation. Plates were centrally inoculated with 5 mm diameter fungal mycelial discs and incubated at 28 °C for 7 days. After incubation, the plates were flooded with Gram’s iodine solution for 5 min and rinsed gently with distilled water. The formation of clear hydrolysis zones surrounding the fungal colonies against a dark blue background was recorded as a positive indication of extracellular amylase activity.
Quantitative enzyme production:
The quantitative enzyme production was carried out using
submerged fermentation in an agro-residue-based medium
[15] with slight modifications. One hundred millilitres of distilled water
was dispensed into 250 mL Erlenmeyer flasks and sterilised by
autoclaving at 121 °C for 15 min. After cooling to room temperature
under aseptic conditions, healthy pomegranate peels (10%, w/v),
thoroughly washed with double-distilled water and surface sterilised
with 70% ethanol, were aseptically added to each flask to serve as
the sole carbon source. The pH of the medium was adjusted to 7.0.
Mycelial discs (5 mm diameter) obtained from a 7-day-old potato
dextrose agar culture of P. arenariae were used as inoculum. The
inoculated flasks were incubated at 28 °C on an orbital shaker at 120
rpm. Uninoculated flasks served as controls. Samples were collected
at 7, 14, 21, and 28 days to evaluate enzyme production dynamics.Preparation of crude enzyme extract:
At each sampling interval, cultures were filtered through
Whatman No. 1 filter paper to remove mycelial biomass. The filtrate
was centrifuged at 10,000 × g for 15 min at 4 °C, and the resulting
supernatant was used as the crude enzyme extract for subsequent
assays.Protein determination:
Total extracellular protein concentration was determined by the
Lowry protein assay using bovine serum albumin as a standard. 1.0
ml of appropriately diluted crude supernatant and 5ml of alkaline
copper reagent were incubated for 10 min, followed by the addition
of diluted Folin–Ciocalteu reagent. The absorbance was measured at
660 nm using a UV–VIS spectrophotometer after incubation in the
dark for 20 min at room temperature. Protein concentration was
calculated from a BSA standard curve and expressed as mg/100 ml
[16].Enzyme assays:
The crude enzyme extract was used for the quantitative estimation
of cellulase, β-glucosidase, pectinase, xylanase, protease, and laccase
activities. The production of these enzymes is associated with the
degradation of plant cell wall components, particularly cellulose,
hemicellulose, and pectin [17,18].
Cellulase: Cellulase activity was determined using 1% (w/v)
carboxymethyl cellulose (HiMedia Laboratories, Mumbai, India)
in 50 mM citrate buffer (pH 5.0). The reaction mixture (1 mL
substrate + 1 mL enzyme) was incubated at 50 °C for 30 min. The
reaction was terminated by adding 2 mL dinitrosalicylic acid (DNS)
reagent prepared according to Miller (1959), followed by boiling
for 5 min. Absorbance was measured at 540 nm using a UV–VIS
spectrophotometer. Reducing sugars were quantified using a glucose
standard curve. Appropriate enzyme and substrate blanks were
included. One unit (U) of cellulase activity was defined as the amount
of enzyme liberating 1 μmol glucose min-¹, expressed as U mL-¹
[19,20].β-Glucosidase: β-Glucosidase activity was assayed using 5 mM p-nitrophenyl-β-D-glucopyranoside in 50 mM phosphate–citrate buffer (pH 5.0). The reaction mixture (1.5 mL substrate + 0.5 mL enzyme) was incubated at 35 °C for 30 min and terminated by adding 1 mL of 1 M Na₂CO₃. Absorbance was measured at 405 nm. The released p-nitro phenol was quantified using a standard curve. Blanks were included for correction. One unit (U) was defined as 1 μmole pNP released min-¹ and expressed as U mL-¹ [21].
Xylanase: Xylanase activity was measured using 1% (w/v) beechwood xylan (TCI Chemicals, Tokyo, Japan) in 50 mM sodium phosphate buffer (pH 5.0). The reaction mixture (1.5 mL substrate + 0.5 mL enzyme) was incubated at 45 °C for 15 min. The reaction was stopped with 2 mL DNS reagent (Miller, 1959) and boiled for 5 min. Absorbance was recorded at 540 nm. Reducing sugars were quantified using xylose standard. One unit (U) corresponded to 1 μmol xylose released min-¹ and was expressed as U mL-¹ [22,23].
Pectinase (polygalacturonase): Pectinase activity was determined using 0.5% (w/v) citrus pectin (HiMedia Laboratories, Mumbai, India) in 50 mM sodium acetate buffer (pH 5.5). The reaction mixture (1 mL substrate + 1 mL enzyme) was incubated at 50 °C for 30 min. The reaction was terminated with 2 mL DNS reagent (Miller, 1959) and boiled for 10 min. Absorbance was measured at 540 nm. Galacturonic acid was used as a standard. One unit (U) was defined as 1 μmol product released min-¹ and expressed as U mL-¹ [23,24].
Protease: Protease activity was assayed using 0.65% casein as
substrate. The reaction mixture was incubated at 37 °C for 30 min
and terminated with trichloroacetic acid. The soluble peptides were
quantified using the Folin–Ciocalteu method, and absorbance was
measured at 660 nm. Tyrosine was used as standard. One unit (U)
of protease activity was defined as 1 μmol tyrosine released min-¹ and
expressed as U mL-¹ [25].
Laccase: Laccase activity was determined using 1 mM ABTS
(Sigma-Aldrich, St. Louis, USA) in 100 mM sodium acetate buffer
(pH 4.5). The reaction mixture (0.9 mL substrate + 0.1 mL enzyme)
was monitored at 420 nm. Activity was calculated using an extinction
coefficient (ε₄₂₀ = 36,000 M-¹ cm-¹) assuming a 1 cm path length. One
unit (U) was defined as the amount of enzyme oxidising 1 μmol ABTS
min-¹ and expressed as U mL-¹ [26].
Fractionation and dialysis of extracellular proteins:
Extracellular proteins were partially purified from the culture
filtrate obtained at day 14 of incubation, corresponding to peak
enzyme production. The culture broth was filtered through Whatman
No. 1 filter paper and centrifuged at 10,000 × g for 15 min at 4 °C to
obtain a clear supernatant. Protein precipitation
[27] was carried out
by gradual addition of solid ammonium sulfate to the supernatant
under continuous stirring at 4 °C to achieve 80% saturation,
following standard salting-out procedures for protein. The mixture
was stirred for 3 h and further incubated overnight at 4 °C to ensure
complete precipitation of extracellular proteins. The precipitated
proteins were recovered by centrifugation at 10,000 × g for 20 min
at 4 °C. The resulting pellet was resuspended in a minimal volume of
50 mM sodium phosphate buffer (pH 7.0) and subjected to dialysis
against the same buffer at 4 °C for 24 h with periodic buffer changes to
remove residual ammonium sulfate, as described in standard protein
purification protocols [28]. The dialysed protein extract was used for
SDS–PAGE analysis.SDS–PAGE analysis of extracellular proteins:
Extracellular proteins were analysed by sodium dodecyl sulfate–
polyacrylamide gel electrophoresis (SDS–PAGE) using a Mini-
PROTEAN Tetra Cell, 2-Gel System, vertical electrophoresis system
(Bio-Rad Laboratories, China), following the method of Ulrich K.
Laemmli (1970) with minor modifications [29]. A discontinuous
gel system consisting of 12% resolving gel and 4% stacking gel
was employed, as described in standard protein electrophoresis
protocols [29,30]. The 12% resolving gel (10 mL) was prepared
by mixing 4.0 mL of 30% acrylamide solution, 2.5 mL of 1.5 M
Tris–HCl buffer (pH 8.8), 0.1 mL of 10% (w/v) SDS, and 3.35 mL
of distilled water. Polymerisation was initiated by adding 50μL
of 10% ammonium persulfate (APS) and 15μL of N, N, N′, N′-
tetramethylethylenediamine (TEMED). The 4% stacking gel (5 mL)
was prepared using 0.665 mL of 30% acrylamide/bis-acrylamide
solution, 1.25 mL of 0.5 M Tris–HCl buffer (pH 6.8), 50μL of 10%
(w/v) SDS, and 3.0 mL distilled water, followed by the addition of
25μL APS and 10μL TEMED to initiate polymerisation. The resolving
gel was cast between glass plates and overlaid with n-butanol to ensure
a uniform surface. After polymerisation, the overlay was removed,
and the stacking gel was poured over the resolving gel. A comb
was inserted to form wells, and the gel was allowed to polymerise
completely. Following polymerisation, the comb was removed and
the wells were rinsed with running buffer. Dialysed protein extract
was loaded, and electrophoresis was carried out at a constant voltage
of 150 V for approximately 50 min, A pre-stained protein molecular
weight marker (MBT092, HiMedia Laboratories, India) was used as
a standard. The apparent molecular weights of protein bands were
estimated by comparing their relative mobility (Rf values) with those
of the marker proteins (Ulrich K. Laemmli, 1970).Statistical analysis:
All experiments were performed in triplicate, and the results are
expressed as mean ± standard deviation (SD).Results
Qualitative profiling of extracellular enzyme activities:
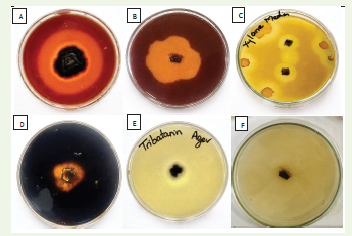
The fungal isolate was screened for extracellular enzyme
production using substrate-specific plate assays. The isolate exhibited
positive activity for cellulase, pectinase, xylanase, amylase, and lipase,
as indicated by the formation of distinct clear halo zones around
the colonies on respective agar media [Figure 1]. The appearance of
these zones confirmed the extracellular hydrolysis of cellulose, pectin,
xylan, starch, and lipid substrates. However, no distinct qualitative
halo zones were observed for protease and laccase, suggesting low
extracellular expression under plate assay conditions; however, low
but measurable activities were detected in submerged fermentation
assays. Overall, the fungal isolate demonstrated strong qualitative
production of multiple extracellular hydrolytic enzymes, except
protease and laccase, during preliminary screening.Time-course quantitative production of extracellular cell wall-degrading enzymes by Paradendryphiella arenariae:
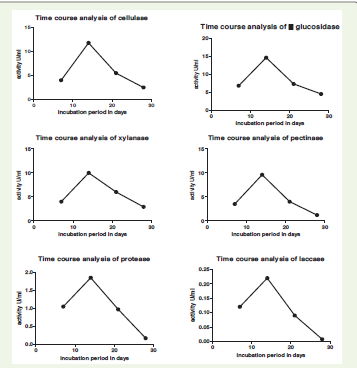
The temporal production of extracellular enzymes by P. arenariae
was evaluated at 7-day intervals up to 28 days under submerged
fermentation [Table 1]. Enzyme activities increased from day 7 to
day 14, followed by a gradual decline. Maximum production of all
enzymes was recorded on day 14 [Figure 2], indicating this as the
optimal incubation period. β-Glucosidase showed the highest activity
(14.6 U/mL), followed by cellulase (11.8 U/mL), xylanase (10.0 U/
mL), and pectinase (9.6 U/mL), demonstrating the pronounced
lignocellulolytic potential of the isolate. The concurrent peak of these
enzymes suggests a coordinated role in lignocellulose degradation,
where cellulases and xylanases hydrolyse structural polysaccharides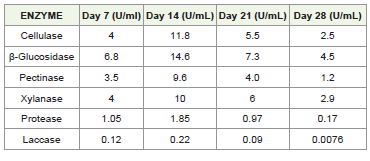
and β-glucosidase facilitates glucose release, enhancing
saccharification efficiency. Such elevated hydrolytic enzyme activity
is also indicative of the organism’s capacity to degrade plant cell wall
components, thereby contributing to tissue maceration and fruit rot
during spoilage. In contrast, protease and laccase were produced at
lower levels, with maximum activities of 1.85 U/mL and 0.22 U/mL,
respectively. A progressive decline in enzyme activities was observed
after day 14, with substantial reductions by day 28, particularly
for laccase (0.0076 U/mL) and protease (0.17 U/mL). Overall, the
integrated CWDE profile supports the pathogenic competence
of P. arenariae, demonstrating its ability to produce a spectrum of
hydrolytic enzymes essential for host cell wall degradation.
Quantification of enzymes:
The total extracellular protein concentration of the culture
filtrate at day 14 was estimated to be 1.61 mg mL-¹. Specific activities
of extracellular enzymes were calculated by normalising enzyme
activity against total protein concentration [Figure 3]. Among the
enzymes, β-glucosidase exhibited the highest specific activity (9.07
U mg-¹ protein), followed by cellulase (7.33 U mg-¹), xylanase (6.21
U mg-¹), and pectinase (5.96 U mg-¹). Protease and laccase showed
comparatively lower specific activities of 1.15 and 0.14 U mg-¹
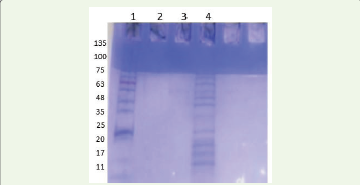
protein, respectively.SDS–PAGE analysis of extracellular proteins:
SDS–PAGE analysis of dialysed crude extracellular enzyme
extracts revealed multiple protein bands ranging from approximately
17 to 75 kDa [Figure 4]. The crude sample (lane 4) showed prominent
bands at ~35–48 kDa, along with additional bands at ~25 kDa and
~17 kDa. The observed banding pattern indicates the presence of
a complex mixture of cell wall–degrading enzymes, with major
proteins corresponding to typical molecular weights of cellulases,
pectinases, β-glucosidases, and xylanases. The intensity of bands in
the mid molecular weight range suggests high enzyme expression at
the selected incubation period (day 14). However, these assignments
are tentative and based on molecular weight comparison.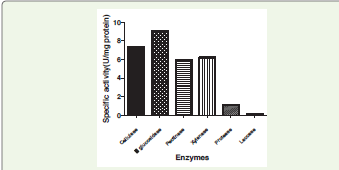
Discussion
The present study provides the first evidence of extracellular cell
wall–degrading enzyme (CWDE) production by Paradendryphiella
arenariae associated with pomegranate fruit rot, supporting its
pathogenic potential and establishing its role as an emerging fungal
pathogen of pomegranate under field conditions. The secretion of
extracellular hydrolytic enzymes is a critical determinant of fungal
pathogenicity, enabling host penetration, tissue maceration, nutrient
acquisition, and disease progression through degradation of structural
polysaccharides in plant cell walls [1-4,6,8].
Qualitative plate assays confirmed the secretion of cellulase, xylanase, pectinase, lipase, and amylase, indicating the broad degradative capacity of P. arenariae. No distinct halo zones were observed for protease and laccase, suggesting comparatively lower expression under plate assay conditions; However, measurable quantitative activities detected under submerged fermentation indicate that enzyme production is influenced by environmental and nutritional factors [15,18].
Time-course of enzyme analysis revealed coordinated induction of cellulase, β-glucosidase, xylanase, and pectinase, with maximum activities recorded on day 14, followed by gradual decline. Such synchronized enzyme production is characteristic of active substrate colonization during fungal growth and reflects efficient adaptation to host-derived lignocellulosic substrates [2,4,18,20]. Among the enzymes, β-glucosidase exhibited the highest activity and specific activity, suggesting efficient cellulose saccharification and sustained fungal carbon utilization. Elevated cellulase and β-glucosidase activities indicate an effective cellulolytic system that likely contributes to tissue softening and structural collapse during infection [20,21]. Substantial pectinase activity observed highlights its likely role in pathogenesis. Pectinases are well-recognized pathogenicity factors in fruit rot fungi due to their ability to hydrolyze pectin-rich middle lamellae, leading to cell separation and tissue maceration [12,24]. Since pomegranate pericarp is rich in pectic polysaccharides, pectinase secretion by P. arenariae likely facilitates early-stage penetration and symptom expansion. Concurrent xylanase production further supports efficient degradation of hemicellulosic cell wall components [10,22].
Protease and laccase activities remained comparatively low throughout incubation, suggesting a secondary role in pathogenicity. Reduced laccase activity may reflect the relatively low lignin content of pomegranate fruit tissues, where extensive oxidative degradation is less critical for host colonization. Similar extracellular enzyme patterns have been reported for fruit-associated necrotrophic fungi, where polysaccharide-degrading enzymes predominate over oxidative enzymes [5,26].
The extracellular protein concentration and corresponding specific activities confirmed active enzyme secretion during peak growth. SDS–PAGE analysis further revealed multiple extracellular proteins ranging from approximately 17–75 kDa, consistent with reported molecular weights of fungal cellulases, xylanases, pectinases, and β-glucosidases [27-30]. The observed protein profile supports the biochemical evidence for a complex hydrolytic secretome involved in host tissue degradation.
Collectively, these findings establish P. arenariae as an emerging pomegranate fruit rot pathogen possessing a robust extracellular hydrolytic arsenal. The coordinated secretion of cellulolytic, pectinolytic, and hemicellulolytic enzymes likely underpins host colonization, symptom development, and rapid fruit decay. These enzymes represent important virulence determinants and potential biochemical targets for developing effective disease management strategies to reduce pre- and postharvest losses in pomegranate [1-4,6,7,18]
Qualitative plate assays confirmed the secretion of cellulase, xylanase, pectinase, lipase, and amylase, indicating the broad degradative capacity of P. arenariae. No distinct halo zones were observed for protease and laccase, suggesting comparatively lower expression under plate assay conditions; However, measurable quantitative activities detected under submerged fermentation indicate that enzyme production is influenced by environmental and nutritional factors [15,18].
Time-course of enzyme analysis revealed coordinated induction of cellulase, β-glucosidase, xylanase, and pectinase, with maximum activities recorded on day 14, followed by gradual decline. Such synchronized enzyme production is characteristic of active substrate colonization during fungal growth and reflects efficient adaptation to host-derived lignocellulosic substrates [2,4,18,20]. Among the enzymes, β-glucosidase exhibited the highest activity and specific activity, suggesting efficient cellulose saccharification and sustained fungal carbon utilization. Elevated cellulase and β-glucosidase activities indicate an effective cellulolytic system that likely contributes to tissue softening and structural collapse during infection [20,21]. Substantial pectinase activity observed highlights its likely role in pathogenesis. Pectinases are well-recognized pathogenicity factors in fruit rot fungi due to their ability to hydrolyze pectin-rich middle lamellae, leading to cell separation and tissue maceration [12,24]. Since pomegranate pericarp is rich in pectic polysaccharides, pectinase secretion by P. arenariae likely facilitates early-stage penetration and symptom expansion. Concurrent xylanase production further supports efficient degradation of hemicellulosic cell wall components [10,22].
Protease and laccase activities remained comparatively low throughout incubation, suggesting a secondary role in pathogenicity. Reduced laccase activity may reflect the relatively low lignin content of pomegranate fruit tissues, where extensive oxidative degradation is less critical for host colonization. Similar extracellular enzyme patterns have been reported for fruit-associated necrotrophic fungi, where polysaccharide-degrading enzymes predominate over oxidative enzymes [5,26].
The extracellular protein concentration and corresponding specific activities confirmed active enzyme secretion during peak growth. SDS–PAGE analysis further revealed multiple extracellular proteins ranging from approximately 17–75 kDa, consistent with reported molecular weights of fungal cellulases, xylanases, pectinases, and β-glucosidases [27-30]. The observed protein profile supports the biochemical evidence for a complex hydrolytic secretome involved in host tissue degradation.
Collectively, these findings establish P. arenariae as an emerging pomegranate fruit rot pathogen possessing a robust extracellular hydrolytic arsenal. The coordinated secretion of cellulolytic, pectinolytic, and hemicellulolytic enzymes likely underpins host colonization, symptom development, and rapid fruit decay. These enzymes represent important virulence determinants and potential biochemical targets for developing effective disease management strategies to reduce pre- and postharvest losses in pomegranate [1-4,6,7,18]
References
5. Baldrian P (2006) Fungal laccases: Occurrence and properties. FEMS Microbiology Reviews 30: 215-242.
Citation
Krishnamurthi AV, Karigar CS. Role of Cell Wall–Degrading Enzymes of Paradendryphiella arenariae, a Novel Pathogen in Fruit Rot Disease of Pomegranate. J Plant Sci Res. 2026;13(1): 295.