Research Article
Physio-Biochemical Responses During Seed Desiccation in Elaeocarpus tuberculatus Roxb.
Babu KP1, Krishnan PN2, Raveendran M3 and Smitha RB4
1Govt. Higher Secondary School, Chittariparamba, Kannur, Kerala, India
2Director, Biodiversity and Ecology, CISSA,Thiruvanathapuram, Kerala, India
3J N Tropical Botanic Garden and Research Institute, Palode, Thiruvanathapuram, Kerala, India
4St Dominic’s College, Kanjirappalli, Kerala, India
2Director, Biodiversity and Ecology, CISSA,Thiruvanathapuram, Kerala, India
3J N Tropical Botanic Garden and Research Institute, Palode, Thiruvanathapuram, Kerala, India
4St Dominic’s College, Kanjirappalli, Kerala, India
*Corresponding author:Krishnan PN, Director, Biodiversity and Ecology, CISSA,Thiruvanathapuram, Kerala, India Email: peringattulli@gmail.com
Copyright: ©Babu KP, et al. 2026. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Article Information:Submission: 12/04/2026; Accepted: 06/05/2026; Published: 09/05/2026
Abstract
Understanding the physio-biochemical responses in Elaeocarpus tuberculatus Roxb. (often called the Wild Rudraksha) involves looking at its behaviour as a recalcitrant (desiccation-sensitive) seed. Unlike common garden seeds that can be dried and stored for years, these seeds are native to moist, tropical environments (like the Myristica swamps of the Western Ghats) and die when they lose much water. The most critical change is the drop in moisture content
during desiccation in E. tuberculatus, the seed loses its ability to germinate. As water leaves the cells, the phospholipid bilayer of the cell membranes becomes unstable. This leads to electrolyte leakage, where essential minerals and salts seep out of the seed resulting in the loss of viability. The lack of water triggers the formation of Reactive Oxygen Species (ROS). These “free radicals” attack the fats (lipids) in the cell membranes, creating malondialdehyde
(MDA). High MDA levels are a primary marker of seed death. Free radical scavenging antioxidant enzymes like peroxidase (PO) and polyphenol oxidase (POD) show a sharp decrease in activity, leaving the seeds vulnerable to oxidative stress. The results of the present study suggests that the lack of raffinosemediated vitrification and uncontrolled oxidative stress are the primary drivers of recalcitrant nature of seeds in E. tuberculatus. These findings provide a vital physiological framework for the ex-situ conservation and cryopreservation of this endemic species.
Keywords:Elaeocarpus Tuberculatus Roxb; Seed Viability; Desiccation; Physiology; Biochemistry
Introduction
The Western Ghats of India, one of the World’s hotspots of
biodiversity is characterised by altitudinal, as well as latitudinal
gradients with tropical climatic factors. The environmental
heterogeneity makes this area an extremely heterogeneous
biogeographic zone, with a large amount of plant and animal
diversity and support a high degree of endemism (30%) having
4500 higher plant species [1]. Many of the trees of this biome are
evolutionarily primitive as far as their morphological, anatomical or
physiological parameters are considered. The main propagule of these
trees are large sized seeds that generally prefer riparian climate for
the successful sustenance of genetic diversity. Large sized seeds, high
moisture content, carbohydrate as predominant reserve food and
vascularisation of the integuments are considered as primitive seed
characters [2].
Elaeocarpus tuberculatus Roxb. (Elaeocarpaceae) commonly
known as the Wild Rudraksha, is a prominent canopy tree species
restricted to the evergreen and Shola forests of the Western Ghats,
India reaching a height of 20-30 meters [3]. The species serves
as a vital keystone component in riparian and Myristica swamp
ecosystems. Additionally, the plant is noted for its medicinal
properties and highly ornamented seeds. Flowers are white in colour.
The fruit is drupe and contain a single seed with two halves. Despite
its ecological and economic importance, the natural regeneration
of this species is severely hindered by the recalcitrant nature of its
seeds. Another important characteristic of this species is the seed
years, i.e. the species exhibits masting habit of 2-5 years. The edaphic
factors of their habitat also have a marked effect on the production
of seeds. Unlike orthodox seeds that undergo maturation drying to
achieve a quiescent state, seeds of E. tuberculatus are shed at high
moisture contents and remain metabolically active. Any reduction in
their critical moisture threshold triggers a sequence of deteriorative
physio-biochemical changes, including membrane disruption,
electrolyte leakage, and the accumulation of reactive oxygen species
(ROS). High vulnerability to viability loss, predation of the seeds and
the poor seed years will definitely pose threat to most of the endemic
taxa of the mega diversity spot of Western Ghats. This study aims to
investigate these changes to identify the specific metabolic ‘tipping
points’ that lead to viability loss giving emphasis to the changes in
sugars and anti-oxidant enzymes that will provide vital data for
the ex-situ conservation and long-term storage of this ecologically
sensitive species.
Materials and Methods
Fruit/seeds of Elaeocarpus tuberculatus were harvested from five
healthy selected matured trees from Ponmudi hills, Trivandrum,
Kerala (8°45’and 8°47’N latitude and 77°1’and 77°4’ E longitude) at
an altitude of 700m. The location experiences a mean temperature
of 16-30°C, relative humidity in the range of 60-90%, light intensity
of 2000-20000 lux/m2 with an average rainfall of 2500-3000mm/
year. To avoid damage and soil borne pathogens, mature fruits were
collected by gently shaking of fruit bearing branches. Collected
fruits were brought to the laboratory in polyethylene bags, washed
and spread on absorbent paper to check fungal and other microbial
attack before de-husking.
Desiccation and viability assessment studies:
Seeds were spread in the laboratory conditions to determine
the change in moisture content and germination during desiccation
on air drying. The whole seeds were sampled at regular intervals
(48 hr) from 0 to 18 days after harvest for the analysis of moisture
content. Another set of seeds in triplicates was used for germination
test. Before sowing for germination, the seeds were transferred to
humidity chamber in order to reduce the imbibitional damage while
transferring directly into the water. Moisture content (MC) of the
seeds at each interval was determined according to ISTA rules [4],
High Constant Air Owen method, i.e., by drying at 130±1°C for 1
hour and was calculated on fresh weight basis and expressed as
percentage of moisture (MC).For estimation of Seed vigour index, ten seeds each in triplicates were taken from the seed lots during different desiccation periods at respective intervals (48 hr from 0 to 18 days after harvest) and sown in garden pots filled with clean sand and kept in the shady area for seed germination. The number of seeds germinated on each day was noted and the seed vigour index (SVI) was calculated according to the formula given by [5].
Tetrazolium reduction activity assay
Dehydrating seeds collected at specific intervals mentioned
elsewhere after cutting into pieces were incubated in 1% 2, 3,
5-triphenyl tetrazolium chloride (in 10 mM sodium phosphate buffer
pH 7.2) in darkness for two hours. After three washes in distilled
water, 5 ml acetone was added and incubated at room temperature
in darkness for 16-18 hours to elute the red colour formazan formed.
Optical density of the elute was measured at 480 nm in Systronics
model 106 spectrophotometer and expressed as A 480 nm/g.fwt.
Effect of desiccation in seeds were evaluated by measuring electrolyte leakage and lipid peroxidation. Five seeds in triplicate from different desiccation period were soaked in 25 ml distilled water after chopping into pieces and covered with a lid to reduce evaporation during incubation and also to check dust contamination. After 24 hours the electrical conductivity of the leachate was measured using a dip cell conductivity meter (Systronics DDR 306). The specific conductance was expressed as μscm-1g.
Effect of desiccation in seeds were evaluated by measuring electrolyte leakage and lipid peroxidation. Five seeds in triplicate from different desiccation period were soaked in 25 ml distilled water after chopping into pieces and covered with a lid to reduce evaporation during incubation and also to check dust contamination. After 24 hours the electrical conductivity of the leachate was measured using a dip cell conductivity meter (Systronics DDR 306). The specific conductance was expressed as μscm-1g.
Lipid peroxidation was measured by measuring the
concentration of thio barbituric acid reactive substances (TBRS),
equated with malondialdehyde (MDA) [6]. One-gram fresh tissue
was homogenized in 1 ml distilled water with sterilized silica in a
mortar and pestle. 2 ml of 5% thiobarbituric acid (TBA) prepared in
20 % trichloroacetic acid was added to the homogenate. The reaction
mixture was incubated in a boiling water bath for 30 minutes. A Pink
colour was developed by the reaction of various lipid peroxidation
products (viz. MDA) with TBA. The test tubes were immediately
transferred to 00C in a freezer to stop the reaction. The colour of the
supernatant after centrifugation (at 3000 rpm for 15 minutes) was
measured at 540 nm and 600nm respectively. After subtracting the
absorbance of the non-specific (at 540 nm) the net absorbance was
expressed at 540 nm mgg-1dr.wt. of the seeds.
Biochemical analysis:
Metabolites including total soluble sugars (TSS), starch, phenol,
protein, lipids and individual sugars (glucose, galactose, arabinose,
raffinose, and sucrose) and the activity of enzymes like peroxidase,
polyphenol oxidase were also analysed for studying the stress induced
changes occurring during the desiccation of the seeds. All operations
except enzyme assay were carried out immediately after moisture
analysis. For enzyme assay the de-coated seeds were soaked in water
before assay.Extraction and estimation of metabolites:
One gm tissue of sample was ground in known volume of 80%
ethanol (v/v) in distilled water and centrifuged at 4000 rpm for 10
minutes. The residue was again washed thrice with ethanol and
after centrifugation the supernatant was collected and made up to
10ml with 80% ethanol. The supernatant served as source for the
quantification of total sugars, phenolics and amino acids. Five ml
of the extract was kept in a china dish and kept in hot air oven at
600C and was dissolved in 5 ml distilled water and was centrifuged at
3000 rpm for 10 minutes and the supernatant served as the source for
soluble sugar estimation. The left-over residue was ground with 4 ml
30% Perchloric acid (PCA), centrifuged at 3000 rpm for 10 minutes
each and the residue was re-extracted with 15% PCA. The combined
supernatant was used for the starch estimation.Total soluble sugar (TSS) content was estimated by using the
method proposed by Montgomery [7]. Total soluble sugar was
calculated using glucose standard graph and expressed as mg g-1dr.wt.
Content of starch in tissues was estimated according to Mc Cready et
al. [8]. The starch content was calculated by multiplying the value with
a constant value of 0.9 and expressed as mg -1g dw using D-Glucose
as the standard. Phenolic content of the tissues was estimated by
following the method of Swain and Hillis [9] and was calculated from
standard graph drawn using catechol as the standard and expressed
as mg g-1dr.wt. Total protein content in E. tuberculatus seeds were
determined using the method of Lowry et al. [10]. Bovine serum
albumin (BSA) was used as standard and the total protein quantity
expressed as mg g-1dr.wt. Total extractable lipids were estimated
using the method of Bligh and Dyer [11]. Total weight of lipids in the
tissues and expressed as mg g-1dr.wt.
Extraction and Estimation of different sugars through HPLC
Sample preparation:
One gram seed sample was soaked in 95% ethanol and crushed in
a mortar to get a fine paste. The mixture was then refluxed for 2 hours
in a water bath. The clear filtrate from the reflux was concentrated in
a rotary evaporator to remove the alcohol. Distilled water was then
added to the mixture to make the volume to 5 ml. Insoluble materials
if any, was filtered off. Lipids were removed by adding petroleum
ether and were separated in a separating funnel. The aqueous extract
thus obtained was filtered through a Whattman No. 1 paper and the
filtrate was used for HPLC analysis [12].HPLC analysis:
The aqueous solutions obtained in the sample preparation were
filtered through a 0.45μm Nylon filters and then passed through
a guard column in HPLC (Shimadzu LC 2010A HT) system.
Acetonitrile-water (70:30) mixture was used as the mobile phase in
the column of LC-NH2 at a flow rate of 1ml/min. 20μl of the sample
was injected into the column for detecting the sugars. Refractive index
detector was used for the detection of sugars. Standard solution of the
sugars like glucose, galactose, arabinose, raffinose, sucrose, glycerol
and ethylene glycol at concentration of 1mg/5ml were injected and
the peak areas were calculated by the automated computer integrator.
Concentrations of the sugars were calculated in comparison with
those of the standard solutions. Retention times for glucose, galactose,
arabinose, raffinose, and sucrose were also noted.Extraction of enzymes:
One-gram fresh seed tissue of each sample was homogenised
in a pre-chilled mortar and pestle with a pinch of purified sand and
0.1 M cold sodium phosphate buffer (pH 7.0). The homogenate was
centrifuged at 5000 rpm for 20 minutes using refrigerated centrifuge
and the supernatant was collected. Proteins from the supernatant
were precipitated in cold acetone twice and re-suspended in the
extraction buffer after the evaporation of the acetone. The extract
was centrifuged and the supernatant served as the enzyme source
for both peroxidase and poly phenol oxidase assay. All the extraction
procedures were carried out at 40C.Peroxidase enzyme assay:
Peroxidase enzyme activity was measured according to the
method of Chance and Maehly [13] by recording the change in
absorbance at 470 nm due to oxidation of guaiacol in presence of
hydrogen peroxide. The change in absorbance was recorded in
spectrophotometer (Systronics model 106) at 470 nm. The enzyme
activity was measured as change in absorbance per minutes using
guaiacol as substrate.Polyphenol oxidase:
The polyphenol oxidase activity was measured by following
the method of Yamaguchi et al. [14]. The assay mixture consisted
of 1.0 ml 0.01M catechol and 1.0 ml 0.1M proline in 2.0 ml 0.1M
Sodium phosphate buffer (pH 6.8). 1.0 ml enzyme extract was added
quickly and the kinetics of the enzyme was recorded at 525 nm in
spectrophotometer (Systronics 106) and the enzyme activity was
expressed as change in absorbance per minute.Statistical analysis:
The data obtained from the experiments were analysed following
Analysis of Variance (ANOVA) and the ratio obtained were checked
for significance at 5% level. The means of each treatment were
separated following the Least Significance Difference (LSD) by
Duncan’s Multiple Range Test (DMRT) at 5% level.Results
Elaeocarpus tuberculatus belongs to typical evergreen canopy
species and are restricted to cool, humid riparian environment. The
fruit of E. tuberculatus is a drupe with a fleshy pericarp and matures
approximately 150 days after anthesis (DAA). The fruit/seed dispersal
is through the agency of frugivores, especially monkeys, bats and
giant squirrels. Natural regeneration is hindered by several biotic and
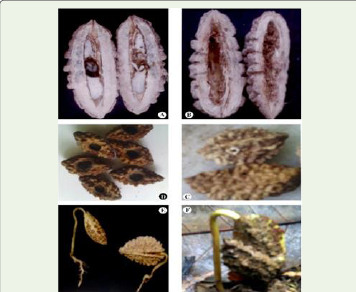
abiotic factors. A large number of beetles are found as seed predators
in the field and also under storage of the seeds (Plate 2). Some of them
make holes in the seed coat and find their way in to the seed locule.
Large number of larvae of insects is found inside the fruit during
different stages of fruit development. Most of them lay eggs when the
fruits are young. A large number of young fruits with holes are found
lying beneath the mother trees [Figure 8] (A, B, C and D).
Effect of desiccation on Moisture content and germination:
Freshly harvested seeds of E. tuberculatus possessed 49.21% MC
with 91.67 % germination during the span of 23 days [Figure 1].
When the seeds are exposed to open laboratory conditions (28±20C
& 65 % RH), no significant change in moisture and germination
percentage was observed up to the 4th day. Subsequently a gradual
decrease in MC and germination percentage was recorded. On the
12th day MC reduced to 30.77% with 58% reduction in germination
percentage compared to the control seeds. As the desiccation reached
the 16th day, seeds have only 19.61% MC with only 20% germination
and complete viability loss in E. tuberculatus seeds was recorded at
17.79 % MC.Effect of desiccation on Seed vigour index:
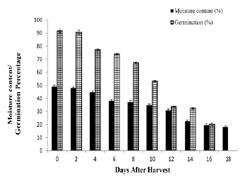
Increase in desiccation period affected the time taken for
germination of the seeds thereby the seed vigour index (SVI) also
decreased accordingly. SVI value of E. tuberculatus was always less than
1 [Figure 2], since the seeds took more than 20 days for germination.
Fresh seeds took 23 days to register 90±2.5 % germination. Their seed
vigour index value was 0.55. Index value decreased gradually from
an initial value of 0.55 to 0.07 on the 12th day. Even though mean
germination days increased gradually from the 12th day onwards there
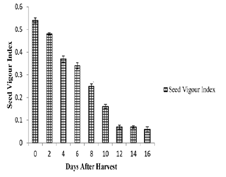
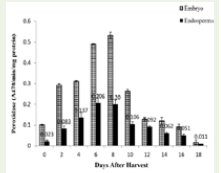
was no change in vigour index and attained a constant value of 0.07.Effect of desiccation on tetrazolium staining pattern and formazan concentration:
Studies on the tetrazolium staining pattern and the colour
development on the embryo showed marked variation during
different desiccation periods. Embryos of fresh seeds showed uniform
intense red colour with TZ. During the successive days of desiccation,
staining pattern and the intensity of staining reduced. On the 6th day
onwards margins of the cotyledons become unstained and the colour
became rose red. By the 12th day the cotyledonary margins become
white in colour and the embryonal axis was rose red. On the 16th day
the axis was feebly stained and no staining on the cotyledons [Table 1].The measurement of formazan concentration was in agreement with the moisture loss and tetrazolium staining pattern. The decreasing trend in the absorbance of the formed formazan was found in E. tuberculatus. On the 12th day onwards the formazan concentration decreased rapidly [Figure 3].
Effect of desiccation on leachate conductivity:
Colour of the seed leachate was yellow in all the samples studied.
More than two-fold increase in specific conductance has been
recorded during the desiccation. Specific conductance of the leachate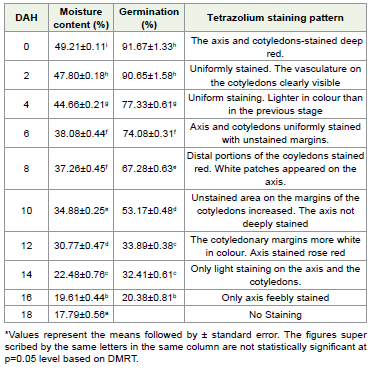
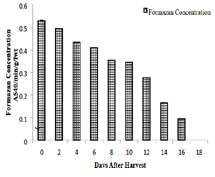
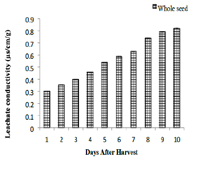
from the desiccating seeds showed an increasing trend. Fresh seeds
registered a value 0.301μs specific conductance which rose to 0.791μs
on the 16th day. About 30% increase in specific conductance occurred
between the 2nd and 4th day [Figure 4].
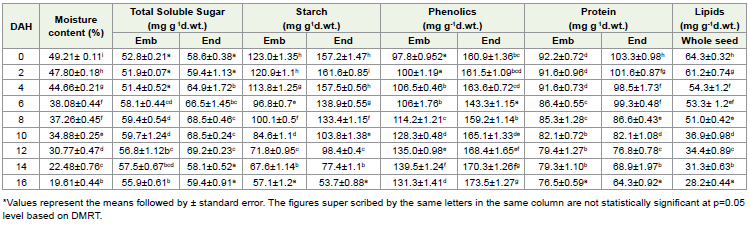
The TSS content in the embryos and endosperm of the fresh seeds was 52.8 ±0.21mg g-1dr.wt. and 58.6±0.38 mg g-1dr.wt. respectively [Table 2]. In embryo the increase in total soluble sugar content was negligible during desiccation without much significant difference [Table 2]. While in the endosperm the TSS content showed an increase up to the 12th day, with a maximum level and later decrease on the 14th and the 16th day of desiccation.
In embryos, starch content was high in fresh seeds (123.0 mg g-1dr. wt. at 49.21% MC), thereafter, starch content decreased gradually as the desiccation continued [Table 2]. Approximately 39% decrease in starch content has been observed during the desiccation. Similar pattern of decreasing trend has been observed in the endosperm also. Highest concentration of starch (161.6 mg g-1dr.wt.) was registered at the 47.80% MC in the endosperm on the 2nd day of which slowly decreased with increase in desiccation.
The TSS content in the embryos and endosperm of the fresh seeds was 52.8 ±0.21mg g-1dr.wt. and 58.6±0.38 mg g-1dr.wt. respectively [Table 2]. In embryo the increase in total soluble sugar content was negligible during desiccation without much significant difference [Table 2]. While in the endosperm the TSS content showed an increase up to the 12th day, with a maximum level and later decrease on the 14th and the 16th day of desiccation.
In embryos, starch content was high in fresh seeds (123.0 mg g-1dr. wt. at 49.21% MC), thereafter, starch content decreased gradually as the desiccation continued [Table 2]. Approximately 39% decrease in starch content has been observed during the desiccation. Similar pattern of decreasing trend has been observed in the endosperm also. Highest concentration of starch (161.6 mg g-1dr.wt.) was registered at the 47.80% MC in the endosperm on the 2nd day of which slowly decreased with increase in desiccation.
In general, higher level of phenolic content has been recorded in
endosperm tissues with an initial content of 160.9 mg g-1dr. wt. [Table 2].
As desiccation increased not much change was noticed. On the
other hand, in embryos the phenolic content was low (156.2±1.2 mg)
with increase in desiccation, the phenolic content also increased with
a maximum of 199.5±0.08 mg g-1dwt-1 on the 14th day.
Analysis of total protein content in the seed of E. tuberculatus revealed that there was a gradual decreasing trend during desiccation in both embryo and endosperm [Table 2]. Negligible difference in protein content has been observed during successive days of desiccation [Table 2].
Total lipid content of the whole seeds of E. tuberculatus on harvest was 64.0 ±0.32 mg g-1dwt, which decreased significantly as the desiccation progressed [Table 2].
Analysis of total protein content in the seed of E. tuberculatus revealed that there was a gradual decreasing trend during desiccation in both embryo and endosperm [Table 2]. Negligible difference in protein content has been observed during successive days of desiccation [Table 2].
Total lipid content of the whole seeds of E. tuberculatus on harvest was 64.0 ±0.32 mg g-1dwt, which decreased significantly as the desiccation progressed [Table 2].
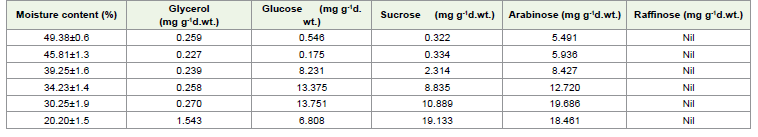
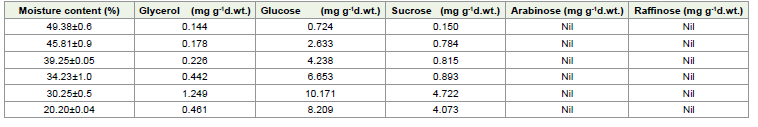
HPLC analysis of simple sugars:
The HPLC analysis revealed the presence as well as increase of
Glycerol, Glucose and sucrose content in the embryos of E. tuberculatus
during desiccation. But arabinose and raffinose were not detected in
any of the samples studied [Table 4]. Of the sugars observed, glucose
was in much higher concentration compared to glycerol and sucrose.
As the moisture content of the seed decreased, the content of these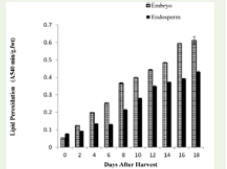
sugars increased. But during the later stages when the moisture
content was 20.20%, the content of these sugars decreased [Table 4]. There was a sharp increase in the levels of these sugars at 30.25%
MC. At this MC, glucose content was 40% higher than the seeds
with 34.25% MC. Similarly, the sucrose content increased abruptly
at this stage. Threefold increase in the concentration of glycerol was
observed when the MC was reduced to 30.25%.
In the endosperm glycerol content did not show much difference up to the 5th day of desiccation (MC 20.20%), which was very high [Table 3], whereas glucose, sucrose, and arabinose recorded a gradual increase from the initial moisture content (49.38%). Interestingly both raffinose and arabinose were not found in the embryos but arabinose was found in the endosperm at a higher concentration. The presence of raffinose was not detected in the seeds of E. tuberculatus.
In the endosperm glycerol content did not show much difference up to the 5th day of desiccation (MC 20.20%), which was very high [Table 3], whereas glucose, sucrose, and arabinose recorded a gradual increase from the initial moisture content (49.38%). Interestingly both raffinose and arabinose were not found in the embryos but arabinose was found in the endosperm at a higher concentration. The presence of raffinose was not detected in the seeds of E. tuberculatus.
Discussion
Desiccation tolerance/sensitivity of seeds is a physiological
phenomenon regulated by a balance between endogenous metabolic
activity as well as exogenous environmental stress. In E. tuberculatus,
the results revealed that there was a positive correlation between
moisture loss and germination percentage [Figure 1]. Fresh seeds
possessed high initial moisture content (49.38%) and registered 90%
germination, and took 23 days to complete germination. When the
seeds exposed to open laboratory conditions (28±20C& 65% RH),
showed a significant decrease in moisture content on the 4th day and
complete loss of viability was recorded on the 18th day. Following
the classification of seeds [15,16,17] E. tuberculatus exhibits typical
,recalcitrant behaviour. The critical moisture content (CMC) defined
by different authors at which 50% viability loss [18] was identified
at an MC of 30.25% in E. tuberculatus. The rapid loss of moisture
content and simultaneous loss of viability as reported in Theobroma
cacao [19], Aporusa lindleyana [20] and Vateria indica [21-23]. The moisture content in the seeds at the time of harvest in tropical and
temperate trees lies between 40% to 60% and for avoiding desiccation
damage maintaining high moisture content is essential [24-26]. Such
type of decrease in moisture content and viability was shown by some
other recalcitrant seeds like Shorea robusta [26], Avicennia marina
[27], Myristica malabarica [28], Mesua ferrea [22] etc. The Seed Vigour Index (SVI) proved to be a sensitive marker
for desiccation-induced deterioration on germination. The low SVI
(0.55) and the prolonged germination period (23 days) in fresh seeds
suggest inherent constraints on rapid emergence. This may be because
of: a. Physical Obstruction due to the thick, highly lignified seed coat
that likely imposes mechanical resistance to radicle protrusion and b.
Chemical Inhibition due to high phenolic concentrations, as observed,
may act as metabolic inhibitors, a phenomenon documented in other
recalcitrant taxa [29]. As desiccation progresses, the drastic decline in
SVI to 0.07 reflects an irreversible collapse of metabolic vigour long
before total embryo death occurs.
At high hydration levels, cell membranes maintain a liquidcrystalline
state, ensuring selective permeability. Dehydration in E.
tuberculatus triggers a transition to a gel phase, leading to physical
“broaching” of the plasma membrane [30]. The observed twofold
increase in leachate conductivity within 16 days confirms that
viability loss is driven by the irreversible efflux of essential solutes as
reported [31]. This negative correlation between electrolyte leakage
and moisture content is a hallmark of desiccation-sensitive species,
where the cellular machinery fails to maintain structural integrity
under water stress [32,33].
The pivotal point of viability loss is closely linked to lipid
peroxidation. The exponential increase in malondialdehyde (MDA)
levels, particularly in the embryo, suggests that reactive oxygen
species (ROS) leads to an uncontrolled “free radical attack” on
membrane lipids. The embryo’s higher MDA concentration relative
to the endosperm (40% higher) reflects its greater physiological
activity and higher susceptibility to oxidative failure as reported
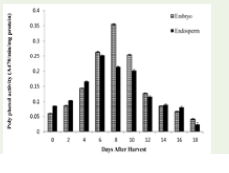
in Cocoa and jack seed [34]. In response to this stress, antioxidant
enzymes like peroxidase (POD) and polyphenol oxidase (PPO)
show altered activity. While POD levels initially increased—likely
as a compensatory mechanism - the subsequent decline at subcritical
moisture levels suggests protein denaturation and metabolic
collapse [35]. The inverse correlation between MDA accumulation
and antioxidant enzyme efficiency is a primary driver of recalcitrant
nature in this species. Similar observations were also made in
Azardirachta indica [36] and in Limonia aureum seeds [37].
The biochemical shift from starch to total soluble sugars (Table 2)
during desiccation suggests active starch hydrolysis or gluconeogenesis
from lipid reserves. According to the water Replacement Theory
[38], sugars like sucrose protect membranes by hydrogen-bonding
to lipid head groups, substituting for water molecules. However, E.
tuberculatus lacks the full “sugar suite” required for true desiccation
tolerance. While sucrose was abundant, the total absence of raffinose
in the embryo is a critical finding. Raffinose-family oligosaccharides
(RFOs) are essential for vitrification (glass formation), which prevents
sucrose from crystallizing and maintains the cytoplasmic matrix in
a stable state [39]. Though the endosperm contained arabinose, its
absence in the vital tissues of the embryo further explains the species’
inability to survive drying.
Conclusion
Elaeocarpus tuberculatus produces recalcitrant seeds whose
survival is strictly bound to high hydration levels. The lethal transition
occurring below 30% MC is driven by lipid peroxidation of the embryo
and a failure in membrane integrity. This will definitely lead to the
failure of all types of anti-oxidant defence system in these seeds. The
lack of raffinose-mediated vitrification and the mechanical barriers
of the seed coat further complicate its natural regeneration. For
conservation purposes, ex-situ strategies must focus on moist storage
or cryopreservation of excised embryos to bypass the limitations of
whole-seed desiccation.
References
7. Montgomery R (1957) Determination of glycogen. Archives of Biochemistry and Biophysics 67: 378-386.
Citation
Babu KP, Krishnan PN, Raveendran M, Smitha RB. Physio-Biochemical Responses During Seed Desiccation in Elaeocarpus tuberculatus Roxb.. J Plant Sci Res. 2026;13(1): 294.