Review Article
The Plant Microbiome: Harnessing Phyllosphere Microbial Communities for Sustainable Cultivation of Sesame (Sesamum indicum)
Prasanna Lakshmi B and Sreeramulu A*
Department of Botany, Sri Venkateswara University, Tirupati, Andhra Pradesh, India
*Corresponding author:Prof. Sreeramulu, Department of Botany, Sri Venkateswara University, Tirupati, Andhra Pradesh, India. E-mail Id: Nandana018@gmail.com
Copyright: © Prasanna Lakshmi B, et al. 2025. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Article Information:Submission: 19/05/2025; Accepted: 05/06/2025; Published: 09/06/2025
Abstract
The phyllosphere microbiome of plants represents a critical, yet underexplored, component of sustainable crop production systems. This review synthesizes current knowledge on the diverse functional roles of leaf-associated microbial communities in enhancing plant productivity and resilience. We examine how phyllosphere microbes contribute to: (1) disease suppression through competitive exclusion, antimicrobial production, and induced systemic resistance; (2) abiotic stress tolerance via biofilm-mediated water retention, ACC deaminase activity, and UVprotective pigmentation; and (3) growth promotion by phytohormone synthesis and nutrient solubilization. Emerging applications in microbiome assisted breeding and precision microbiome management are discussed as innovative approaches to develop climate-resilient varieties. We highlight successful field implementations of microbial consortia that reduce chemical inputs while maintaining yields, including biofertilizer blends and biocontrol formulations effective against major pathogens like Cercosporasesami and Alternaria sesami. Key challenges in mechanistic
understanding, microbial product standardization, and farmer adoption are addressed, along with future directions integrating multi-omics technologies and policy frameworks. The review underscores the phyllosphere microbiome’s potential to transform plant cultivation into a more
productive, sustainable, and climate-smart agricultural system through ecological intensification strategies that harness beneficial plant-microbe interactions. This knowledge provides a foundation for advancing microbiome-based solutions in oilseed crop production globally. The phyllosphere
microbiome of plants represents a critical, yet underexplored, component of sustainable crop production systems. This review synthesizes current knowledge on the diverse functional roles of leaf-associated microbial communities in enhancing plant productivity and resilience. We examine how phyllosphere microbes contribute to: (1) disease suppression through competitive exclusion, antimicrobial production, and induced systemic resistance; (2) abiotic stress tolerance via biofilm-mediated water retention, ACC deaminase activity, and UV-protective pigmentation; and (3) growth promotion by phytohormone synthesis and nutrient solubilization. Emerging applications in microbiome-assisted breeding and precision microbiome management are discussed as innovative approaches to develop climate-resilient varieties. We highlight successful field implementations of microbial consortia that reduce chemical inputs while maintaining yields, including biofertilizer blends and biocontrol formulations effective against major pathogens like Cercosporasesami and Alternaria sesami. Key challenges in mechanistic understanding, microbial product standardization, and farmer adoption are addressed, along with future directions integrating multi-omics technologies and policy frameworks. The review underscores the phyllosphere microbiome’s potential to transform plant cultivation into a more productive, sustainable, and climate-smart agricultural system through ecological intensification strategies that harness beneficial plant-microbe interactions. This knowledge provides a foundation for advancing microbiome-based solutions in oilseed crop production globally.
Keywords:Sesamum Indicum; Phyllo Sphere; Sustainable Agriculture; Plant Growth Promotion; Biocontrol;Stress Tolerance; Microbial Consortia.
Introduction
Sesame (Sesamum indicum L.), a diploid annual plant (Figure
1) belonging to the Pedaliaceae family, stands as one of the oldest
and most economically significant oilseed crops, with a cultivation
history spanning over 5,000 years. Primarily grown for its oil-rich
seeds-containing 45–60% high-quality oil and 18–25% proteinsesame
is a cornerstone of agriculture in tropical and subtropical
regions, particularly in arid and semi-arid zones where its exceptional
drought tolerance makes it indispensable for sustainable farming
systems. Major global producers include India, Sudan, Myanmar,
Tanzania, and Nigeria, where sesame serves as a critical crop for both
local consumption and international trade [1].
Beyond its direct agronomic and nutritional value, sesame is prized
for its nutraceutical and industrial applications. The oil, renowned
for its oxidative stability, is rich in bioactive lignans like sesamin
and sesamolin, which contribute to its widespread use in culinary,
pharmaceutical, and cosmetic industries. Microbial ecosystems
offer eco-friendly alternatives to conventional agrochemicals by
enhancing nutrient uptake, suppressing pathogens, and mitigating
abiotic stresses such as drought, salinity, and heavy metal toxicity. As
global agriculture shifts toward sustainable practices, understanding
and harnessing plant microbiome presents a transformative
opportunity to improve yields, reduce chemical dependency, and
enhance climate resilience-securing sesame’s place as a vital crop for
future food and economic systems [2,3].
Despite its adaptability, sesame production faces
significant challenges due to biotic stresses (such as fungal
pathogens Macrophominaphaseolina causing charcoal
rot, Fusarium spp. inducing wilt, and Cercospora spp. leading to
leaf spot) and abiotic stresses (including drought, salinity, and
heavy metal toxicity) [4,5]. Conventional agricultural practices
often rely on chemical fertilizers and synthetic pesticides to mitigate
these challenges, but their excessive use has led to environmental
degradation, soil health deterioration, and economic burdens for
farmers. Consequently, there is a growing shift toward microbiome based
sustainable agriculture, which leverages the natural microbial
communities to enhance growth, improve stress tolerance, and boost
yield without relying on harmful agrochemicals [6].
Emerging research highlights those plants, hosts a diverse and

dynamic microbiome that plays a crucial role in its survival and
productivity. These microbial communities contribute to Nutrient
cycling (e.g., nitrogen fixation, phosphate solubilization), Disease
suppression (through competition, antibiosis, and induced systemic
resistance) and Stress mitigation (drought tolerance, salinity
adaptation, and heavy metal detoxification). Understanding these
plant-microbe interactions is essential for developing microbiomeenhanced
cultivation strategies that align with global sustainability
goals, such as reducing chemical inputs, improving soil health, and
increasing crop resilience in the face of climate change [7].
This review provides a comprehensive exploration of
1. Microbial diversity in the phyllosphere and rhizosphere – Taxonomic composition, core microbiome, and factors influencing microbial colonization.
2. Functional roles of plant-associated microbes – Mechanisms of plant growth promotion, pathogen inhibition, and stress alleviation.
3. Nutrient acquisition and stress tolerance – How microbes enhance phosphorus uptake, nitrogen fixation, and osmotic regulation under drought and salinity.
4. Agricultural applications – Development of microbial inoculants (biofertilizers, biostimulants, and biocontrol agents) for sesame farming.
5. Research gaps and future directions – Integrating omics technologies, synthetic microbial communities (SynComs), and precision microbiome engineering for optimized sesame production
1. Microbial diversity in the phyllosphere and rhizosphere – Taxonomic composition, core microbiome, and factors influencing microbial colonization.
2. Functional roles of plant-associated microbes – Mechanisms of plant growth promotion, pathogen inhibition, and stress alleviation.
3. Nutrient acquisition and stress tolerance – How microbes enhance phosphorus uptake, nitrogen fixation, and osmotic regulation under drought and salinity.
4. Agricultural applications – Development of microbial inoculants (biofertilizers, biostimulants, and biocontrol agents) for sesame farming.
5. Research gaps and future directions – Integrating omics technologies, synthetic microbial communities (SynComs), and precision microbiome engineering for optimized sesame production
By synthesizing current knowledge on plant microbiome
dynamics, this review aims to bridge the gap between fundamental
research and practical agricultural applications, paving the way
for next-generation, microbiome-driven sesame farming systems.
Geographical distribution, Nutrition profile, phytochemistry and biological functions of Sesame:
S. indicumL., stands as one of humanity’s most ancient and
nutritionally significant oilseed crops, with archaeological evidence
tracing its domestication back to 3500-3050 BCE in the Harappan
civilization. This remarkably resilient plant, belonging to a genus
of approximately 20 species with S. Indicum being the most widely
cultivated, thrives across tropical and subtropical regions in over 60
countries worldwide, with major production centers in India, Sudan,
Myanmar, Tanzania, and China [8]. The small but mighty sesame
seed packs an extraordinary nutritional punch, containing 45-60%
of exceptionally high-quality oil predominantly composed of hearthealthy
polyunsaturated fatty acids (35-50% linoleic acid and 35-50%
oleic acid), along with 18-25% complete protein that includes all
essential amino acids with particularly high levels of methionine - an
amino acid often limited in plant proteins [9]. Beyond its impressive
macronutrient profile, sesame seeds serve as an exceptional source of
dietary minerals, boasting remarkable concentrations of calcium (975
mg/100g), magnesium (351 mg/100g), iron (14.6 mg/100g), and zinc
(7.8 mg/100g), complemented by substantial amounts of B-complex
vitamins and vitamin E in the form of tocopherols, as well as 5-10%
dietary fiber that supports digestive health [10].What truly distinguishes sesame from other oilseeds is its
remarkable array of bioactive phytochemicals that confer numerous
health benefits. The seeds contain unique lignans including sesamin
(0.1-1.1%) and sesamolin (0.1-0.7%), which during processing can
convert to sesamol - compounds that have attracted significant
scientific interest for their potent antioxidant and anti-inflammatory
properties [11]. The phytochemical profile extends to include
valuable phytosterols (β-sitosterol at 300-500 mg/100g, along with
campesterol and stigmasterol), diverse polyphenols such as ferulic
and caffeic acids, aromatic pyrazines like tetramethylpyrazine that
contribute to its distinctive nutty flavor, and lesser-known but equally
important iridoid compounds including sesamoside and verbascoside
[12]. Additionally, sesame contains unique proteins such as globulin
and albumin, which contribute to its functional properties in food
systems and may have additional health benefits. These bioactive
components work synergistically through multiple mechanisms to
provide an impressive spectrum of biological activities that modern
science continues to elucidate [13].
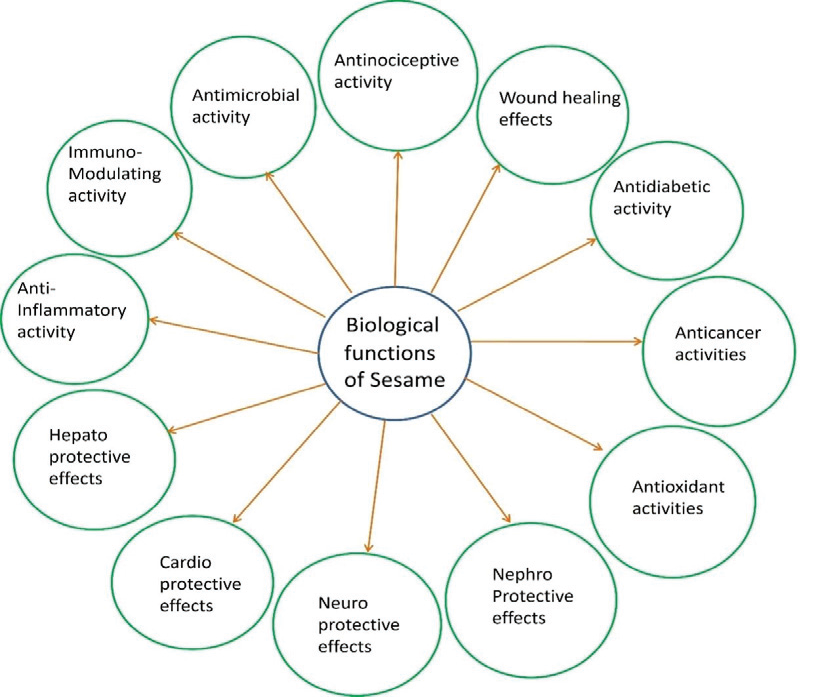
The comprehensive health-promoting properties of sesame are
as diverse as its phytochemical composition. Its potent antioxidant
capacity, primarily attributed to the lignans and tocopherols, operates
through multiple pathways - directly scavenging harmful reactive
oxygen and nitrogen species (ROS/RNS) while simultaneously
enhancing the body’s endogenous antioxidant defence systems
by boosting enzymes like superoxide dismutase (SOD), catalase
(CAT), and glutathione peroxidase (GPx), while also protecting
cellular membranes and DNA from oxidative damage [14,15]. The
cardiovascular benefits of sesame emerge through an impressive
array of mechanisms including notable hypolipidemic effects that
reduce LDL cholesterol and triglycerides, inhibition of cholesterol
absorption in the intestine, improvement of endothelial function,
and antihypertensive activity through ACE inhibition[16]. The
anti-inflammatory potential of sesame compounds manifests
through their ability to modulate critical inflammatory pathways,
particularly by inhibiting NF-κBsignalling, reducing production
of pro-inflammatory cytokines (TNF-α, IL-6), and decreasing
expression of inflammatory enzymes (COX-2 and iNOS) [17].
Emerging research continues to uncover new therapeutic dimensions
of sesame, from hepatoprotective effects that safeguard liver function
through enhancement of hepatic detoxification enzymes, reduction
of lipid accumulation in hepatocytes, and protection against
alcohol- and toxin-induced damage, to neuroprotective properties
that may combat cognitive decline through acetylcholinesterase
inhibition, protection against β-amyloid toxicity, and reduction of
neuroinflammation [18,19]. Current investigations are revealing
promising antidiabetic effects through α-glucosidase and α-amylase
enzyme inhibition, enhanced insulin sensitivity, and pancreatic β-cell
protection, along with anticancer potential demonstrated through
antiproliferative effects on cancer cells, induction of apoptosis, and
inhibition of angiogenesis [20, 21].
This extraordinary combination of nutritional value and
medicinal properties (Figure 2) has cemented sesame’s status across
multiple traditional medicine systems. In Ayurveda, sesame oil is
extensively used for massage therapy (abhyanga) and as a carrier
for herbal preparations [22], while Traditional Chinese Medicine
employs black sesame for liver and kidney toxification [23], and
Middle Eastern medicine utilizes sesame for digestive disorders
[24]. In modern applications, sesame has found important roles
as functional food ingredients, nutraceutical supplements, and in
cosmetic and pharmaceutical formulations [25].
Agriculture perspective and Challenges in Sesame Production: Biotic and Abiotic Stresses:
From an agricultural perspective, sesame is a drought-tolerant
crop, making it crucial for arid and semi-arid farming systems where
water scarcity limits the cultivation of other crops. Its ability to
thrive under low moisture conditions positions it as a key species for
sustainable agriculture in challenging environments [26]. However,
its productivity is often constrained by biotic stresses, such as
fungal pathogens like Macrophominaphaseolina causing charcoal
rot, Fusarium wilt, and Cercospora leaf spot, as well as abiotic stresses
like drought, salinity, and heavy metal toxicity [27]. These stressors
reduce yield and quality, necessitating interventions to enhance
resilience and ensure stable production. Conventional farming
relies heavily on chemical fertilizers and pesticides, which pose
environmental and economic challenges, including soil degradation,
water pollution, and high input costs. This unsustainable reliance
has spurred interest in microbiome-based sustainable agriculture as
a viable alternative. Leveraging the natural microbial communities
associated with plants to enhance growth, stress resilience, and yield
offers a promising solution to reduce agrochemical dependency.
These beneficial microbes can improve nutrient uptake, protect
against pathogens, and help the plant withstand harsh environmental
conditions [28].Emerging research reveals that plantsharbors complex microbial
ecosystems in its phyllosphere (aerial plant parts) and rhizosphere
(root zone), which significantly influence its health and productivity.
These microbial communities form symbiotic relationships with
the plant, contributing to essential physiological and biochemical
processes. These microbial communities contribute to nutrient
cycling, disease suppression, and stress mitigation, offering ecofriendly
alternatives to agrochemicals while promoting long-term
soil health [29]. By harnessing these interactions, farmers can
reduce their reliance on synthetic inputs and adopt more sustainable
practices. Understanding these interactions is critical for developing
microbiome-enhanced sesame cultivation strategies that align with
global sustainable agriculture goals. Such advancements could
revolutionize sesame farming, making it more resilient, productive,
and environmentally friendly in the face of climate change and
resource limitations [30].
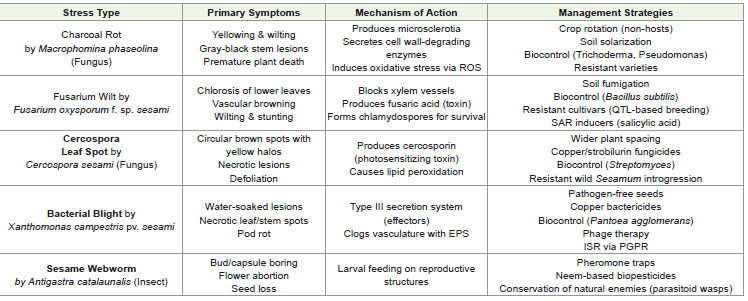
Biotic Stresses: Pathogens and Pests in Sesame Cultivation:
Sesame faces significant threats from a diverse array of fungal,
bacterial, and viral pathogens, as well as insect pests, which
collectively contribute to substantial yield losses and economic
burdens for farmers worldwide. These biotic stressors not only reduce
crop productivity but also compromise seed quality, oil content, and
overall plant vigor. The most devastating diseases include charcoal
rot [31,32], Fusarium wilt [33], Cercospora leaf spot[34]and bacterial
blight[35] each with distinct pathogenic mechanisms and ecological
impacts. Additionally, insect pests such as the sesame webworm
(Antigastracatalaunalis), aphids (Aphis gossypii), and whiteflies
(Bemisiatabaci) cause direct damage through feeding activities while
serving as vectors for viral transmission [36,37].The complexity of
these biotic interactions is further exacerbated by environmental
factors, crop management practices, and the genetic susceptibility of
sesame varieties. Conventional control methods predominantly rely
on chemical fungicides and pesticides, which, while effective in the
short term, pose long-term challenges including pathogen resistance,
environmental contamination, and disruption of beneficial soil
microbiota [31-37].Charcoal Rot (Macrophominaphaseolina) of Sesame:: Charcoal rot, caused by the soil-borne fungus Macrophominaphaseolina, is
one of the most destructive diseases affecting sesame, particularly
in arid and semi-arid regions where high temperatures and drought
conditions favour its proliferation. The pathogen survives in soil and
crop debris as microsclerotia, which germinate under favourable
conditions to infect roots and lower stems (Figure 3). Initial
symptoms include yellowing and wilting of leaves, followed by the

development of grayish-black lesions on stems and roots, resembling
charcoal dust-hence the disease’s name. As the infection progresses,
the fungus colonizes vascular tissues, obstructing water and nutrient
transport and leading to premature plant death. Yield losses can reach
up to 50%, with severe infections resulting in complete crop failure
[31,32]. The pathogen’s virulence is attributed to its production of cell
wall-degrading enzymes (e.g., pectinases and cellulases) and toxins
that disrupt plant cell integrity. Additionally, M. phaseolina induces
oxidative stress in host plants by generating reactive oxygen species
(ROS), which overwhelm the plant’s antioxidant defense systems.
Management strategies include crop rotation with non-host plants,
soil solarization to reduce microsclerotia load, and the application
of biocontrol agents such as Trichoderma spp. and Pseudomonas
fluorescens, which antagonize the pathogen through competition and
antibiosis. Breeding for resistant varieties and maintaining optimal
soil moisture levels are also critical for mitigating charcoal rot’s
impact [38,39].
Fusarium Wilt (Fusarium oxysporumf. sp. sesami) of Sesame:: Fusarium wilt, caused by Fusarium oxysporum f.sp. sesami, is a
vascular disease that poses a significant threat to sesame production,
particularly in regions with warm, moist soils. The pathogen enters
the plant through root tips or wounds, colonizing the xylem vessels
and producing hyphae and spores that obstruct water flow. Early
symptoms include chlorosis of lower leaves, followed by progressive
wilting, stunting, and eventual plant death. The fungus secretes
mycotoxins, such as fusaric acid, which disrupt membrane integrity
and inhibit key enzymatic processes in the host [33].Moreover, F.
Oxysporum induces the formation of tyloses and gels in xylem vessels,
exacerbating water stress. The pathogen’s ability to persist in soil
for years as chlamydospores complicates control efforts. Chemical
treatments are often ineffective due to the pathogen’s protected
vascular niche, necessitating integrated approaches such as soil
fumigation, biocontrol with Bacillus subtilis, and the use of resistant
cultivars [40,41].Recent advances in molecular breeding have
identified quantitative trait loci (QTLs) associated with Fusarium wilt
resistance, offering hope for developing durable genetic solutions.
Additionally, priming plants with salicylic acid or jasmonic acid
can enhance systemic acquired resistance (SAR), reducing disease
severity [33, 40,41].
Cercospora Leaf Spot (Cercosporasesami) of Sesame:
Cercospora leaf spot, caused by the fungus Cercosporasesami,
is a foliar disease that severely impacts sesame photosynthesis and
biomass accumulation. The pathogen spreads via wind-dispersed
conidia, which germinate on leaf surfaces under high humidity and
moderate temperatures. Initial symptoms appear as small, circular,
brownish spots with yellow halos, which coalesce into larger necrotic
lesions, leading to defoliation and reduced photosynthetic capacity.
The fungus produces cercosporin, a photosensitizing toxin that
generates singlet oxygen under light, causing lipid peroxidation
and cell membrane damage in host tissues [34].Severe infections
can reduce yields by up to 30%, with quality losses due to impaired
seed filling. Cultural practices such as wider plant spacing, removal
of infected debris, and avoidance of overhead irrigation can reduce
disease incidence. Fungicidal sprays containing copper-based
compounds or strobilurins are commonly used, but resistance
development necessitates alternation with biocontrol agents like
Streptomyces griseoviridis. Breeding programs are increasingly
focusing on identifying and introgressing Cercospora-resistant traits
from wild Sesamum species into elite cultivars [34,42].Bacterial Blight (Xanthomonas campestrispv. sesami) of Sesame:: Bacterial blight, caused by Xanthomonas campestrispv.
sesami, is a devastating disease characterized by water-soaked lesions
on leaves, stems, and pods, which later turn necrotic and lead to
tissue collapse [35]. The pathogen enters through stomata or wounds,
secreting effector proteins via type III secretion systems to suppress
plant immunity. It also produces extracellular polysaccharides (EPS)
that clog vascular tissues, exacerbating water stress. Warm, humid
conditions favor disease spread, with rain splash and contaminated
tools serving as primary dissemination vectors. Yield losses can
exceed 40% in severe outbreaks. Management includes the use of
pathogen-free seeds, copper-based bactericides, and biocontrol with
Pantoeaagglomerans, which competes for ecological niches. Resistant
varieties are limited, underscoring the need for genomic approaches
to identify resistance genes. Phage therapy and induced systemic
resistance (ISR) via plant growth-promoting rhizobacteria (PGPR)
are emerging as innovative control strategies [35,43].
Insect Pests: Sesame Webworm, Aphids, and Whiteflies:
Insect pests such as the sesame webworm (A. catalaunalis), aphids
(Aphis gossypii), and whiteflies (Bemisiatabaci) cause direct damage
through feeding and indirect harm via virus transmission. The sesame
webworm larvae bore into buds and capsules, leading to flower
abortion and seed loss. Aphids and whiteflies excrete honeydew,
promoting sooty mold growth and reducing photosynthesis. Both
pests transmit devastating viruses like sesame phyllody phytoplasma.
Conventional insecticides are increasingly ineffective due to
resistance, prompting the adoption of IPM strategies. These include
pheromone traps for webworms, neem-based biopesticides, and
conservation of natural enemies like lady beetles and parasitoid wasps
[36,37].(Table 1) summarizes major biotic stresses affecting sesame,
including pathogens (fungal, bacterial) and insect pests, detailing their
causal organisms, characteristic symptoms, mechanisms of infection/
damage, and recommended sustainable management approaches.
ROS (Reactive Oxygen Species), EPS (Extracellular Polysaccharides),
SAR (Systemic Acquired Resistance), ISR (Induced Systemic
Resistance), PGPR (Plant Growth-Promoting Rhizobacteria), QTL
(Quantitative Trait Locus). Management strategies emphasize
integrated approaches combining biocontrol, resistant varieties, and
cultural practices over chemical dependence.Abiotic Stresses: Drought, Salinity, and Heavy Metals in Sesame Cultivation:
Sesame frequently encounters challenging environmental
conditions that significantly impact its growth and productivity.
Among the most critical abiotic stresses are drought, salinity,
and heavy metal toxicity, each posing unique physiological and
biochemical challenges to the plant. These stresses are particularly
prevalent in arid, semi-arid, and industrially affected regions where
sesame is commonly cultivated. While traditional breeding and
genetic engineering have made strides in developing stress-tolerant
varieties, these methods often involve lengthy processes and regulatory
complexities. In contrast, leveraging the plant’s microbiome presents
a sustainable and efficient alternative to enhance stress resilience.
Below, we explore each abiotic stressor in detail, examining their
mechanisms of action and potential mitigation strategies [44,45].Drought Stress in Sesame Cultivation:: Drought stress is a major constraint in sesame cultivation, particularly in regions
with erratic rainfall and high temperatures. Under water-deficient
conditions, sesame plants experience reduced turgor pressure,
leading to stomatal closure and diminished CO2 uptake, which
directly impairs photosynthesis. Prolonged drought also disrupts
nutrient transport, resulting in poor seed filling and yield losses of
up to 60%. At the cellular level, drought induces oxidative stress
through the accumulation of reactive oxygen species (ROS), which
damage lipids, proteins, and DNA [44,45]. To cope, sesame activates
drought-responsive genes and synthesizes osmoprotectants like
proline and glycine betaine, which help maintain cellular hydration.
Microbiome-assisted approaches, such as inoculation with drought tolerant
rhizobacteria (e.g., Azospirillum and Bacillus spp.), enhance
water-use efficiency by improving root architecture and producing
exopolysaccharides that retain soil moisture [46]. Additionally,
agronomic practices like mulching and drip irrigation can mitigate
drought effects, but integrating microbial solutions offers a more
sustainable and cost-effective strategy [44-47].
Salinity Stress in Sesame Cultivation: Salinity stress affects sesame growth by disrupting ion homeostasis and inducing osmotic
and ionic toxicity. High concentrations of sodium (Na+) and
chloride (Cl−) in the soil interfere with potassium (K+) and calcium
(Ca2+) uptake, essential for enzyme activation and cell signalling.
The resulting ionic imbalance impairs metabolic processes, leading
to stunted growth, leaf chlorosis, and reduced oil content in seeds.
Salinity also exacerbates oxidative stress by generating ROS,
which damage cellular membranes and macromolecules. Sesame
plants employ several adaptive mechanisms, including selective
ion exclusion, compartmentalization of toxic ions in vacuoles,
and synthesis of compatible solutes like proline and trehalose.
Microbial interventions, such as the use of halotolerant PGPR
(e.g., Halomonas and Arthrobacter spp.), can alleviate salinity
stress by producing ACC deaminase to reduce ethylene levels,
enhancing antioxidant defenses, and facilitating nutrient uptake. Soil
amendments like gypsum and organic compost also help reclaim
saline soils, but microbiome-based solutions are increasingly favored
for their ecological benefits [48,49].
Heavy Metal Toxicity: Heavy metal contamination, particularly
from cadmium (Cd), lead (Pb), and arsenic (As), poses a severe
threat to sesame cultivation in industrially polluted or wastewaterirrigated
soils. These metals accumulate in plant tissues, disrupting
physiological processes such as photosynthesis, respiration, and
nutrient assimilation. Cd, for instance, replaces Zn in critical
enzymes, rendering them nonfunctional, while lead disrupts cell
division and root elongation. Heavy metals also induce oxidative
stress by catalyzing ROS production, leading to lipid peroxidation and
protein denaturation. Sesame plants employ detoxification strategies
like phytochelatin synthesis, metal sequestration in vacuoles, and
upregulation of antioxidant enzymes (e.g., superoxide dismutase and
catalase). Microbial bioremediation, using metal-tolerant bacteria
(e.g., Pseudomonas and Burkholderia spp.) and mycorrhizal fungi,
offers a promising solution by immobilizing metals in the rhizosphere
or facilitating their uptake and sequestration in non-edible plant parts.
Phytoremediation, coupled with microbial augmentation, can restore
contaminated soils while maintaining crop productivity, though
long-term monitoring is essential to ensure food safety [50,51].
(Table 2) represents summary of major abiotic stresses in sesame
cultivation, including their physiological effects, plant adaptation
mechanisms, and sustainable mitigation strategies. It highlights the
role of microbiome-assisted approaches in enhancing stress resilience
compared to conventional methods.
The Role of plant Microbiomes in Sustainable Cultivation:
Plants are not solitary organisms; they function as holobionts—
complex ecosystems consisting of the host plant and its associated
microbial communities. These microbes form dynamic and evolving
relationships with the plant, significantly influencing sesame growth,
health, and resilience to environmental stresses. By facilitating
nutrient uptake, suppressing diseases, and enhancing drought
tolerance, these microbial partners are indispensable for sustainable
sesame farming [52]. The plant microbiome is shaped by factors such
as soil type, climate, and plant genetics, making it a cornerstone of
crop productivity. Understanding these microbial networks can lead
to innovative cultivation practices that minimize reliance on synthetic
fertilizers and pesticides. These communities primarily reside in the
phyllosphere (aerial plant parts) and rhizosphere (root zone), each
harboring distinct but interconnected microbial ecosystems [53].Phyllosphere Microbiome:
The phyllosphere encompasses the aerial parts of the plant,
including leaves, stems, and flowers, which host diverse microbial
populations. This environment is harsh due to exposure to UV
radiation, temperature fluctuations, and limited nutrient availability.
Despite these challenges, beneficial bacteria, fungi, and yeasts
successfully colonize the phyllosphere, engaging in mutualistic
interactions such as nitrogen fixation, phytohormone production,
and pathogen defense. Some microbes also enhance the plant’s
tolerance to abiotic stresses like heat and drought.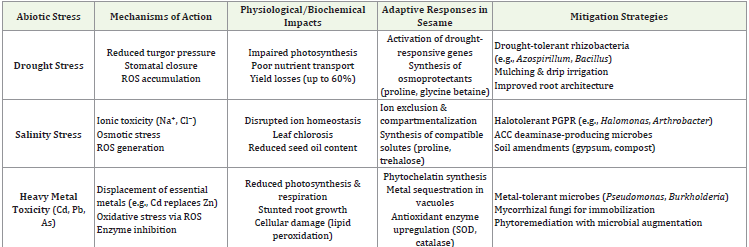
(Figure 4) highlights that the phyllosphere harbours potential
endophytes, which may establish short-term associations or evolve
into long-term mutualistic relationships. These endophytes can be
pathogenic or non-pathogenic, and their colonization dynamics
may displace existing residents. Over time, core spermatophy tea ssociated
microbes and new endophytes contribute to stable,
beneficial partnerships. Research into the phyllosphere microbiome
can unlock natural biocontrol strategies and improve crop resilience
to environmental stressors [54-56].
Rhizosphere Microbiome:
The rhizosphere, the soil region directly influenced by root
exudates, is a hotspot of microbial activity. It hosts a rich consortium
of bacteria (e.g., PGPR—Plant Growth-Promoting Rhizobacteria),
fungi (such as mycorrhizae), and archaea. These microbes play
pivotal roles in nutrient cycling, including phosphorus solubilization
and nitrogen fixation, while also suppressing soil-borne pathogens
through antibiotic production and resource competition.As illustrated in the (Figure 4) the rhizosphere contains potential
endophytes that may colonize seeds or adult plants, contributing
to nutrient uptake and stress tolerance. Like the phyllosphere, the
rhizosphere microbiome includes core microbial endophytes (e.g.,
spermatophytes) and temporal residents, with new arrivals potentially
altering the community structure. These interactions enhance
soil aggregation, water retention, and plant health. Harnessing
rhizosphere microbiomes can lead to sustainable farming practices
that boost yields while reducing chemical inputs [56-58].
The Phyllosphere: A Microbial Hotspot on Leaves and Stems:
The phyllosphere refers to the above-ground surfaces of plants,
including leaves, stems, flowers, and fruits, which serve as dynamic
habitats for diverse microbial communities. These microorganisms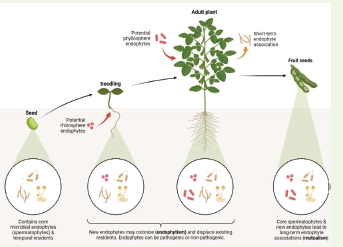
form complex ecological networks that influence plant health, growth,
and stress resilience. Despite harsh conditions-such as intense UV
radiation, temperature fluctuations, and limited nutrient availability the
phyllosphere sustains a rich and active microbiome. These
microbes play crucial roles in nutrient cycling, pathogen defence, and
environmental stress mitigation. Understanding the phyllosphere
microbiome is key to developing sustainable cultivation practices that
enhance crop productivity naturally [59,60].
High-throughput sequencing studies have identified dominant
bacterial phyla, including Proteobacteria (e.g., Pseudomonas and
Methylobacterium), known for their role in nitrogen fixatio¬¬n and
plant growth promotion; Firmicutes (e.g., Bacillus), which enhance
stress tolerance and suppress pathogens; and Actinobacteria (e.g.,
Streptomyces), celebrated for their antibiotic-producing capabilities.
Fungal communities are equally diverse, primarily consistingof
Ascomycota (Alternaria, Cladosporium) and Basidiomycota
(Cryptococcus), w¬¬hich contribute to organic matter decomposition
and symbiotic relationships. Additionally, genera such as Aspergillus,
Trichoderma, and Cladosporium play dual roles—some strains act
as beneficial biocontrol agents, while others may be opportunistic
pathogens under stress conditions. The phyllosphere microbiome
serves as a vital ecological interface, performing multiple functions
that enhance plant fitness, productivity, and resilience. These
microbial communities act as the plant’s first line of defence against
biotic and abiotic stresses while actively promoting growth through
sophisticated biochemical interactions [61-63]..
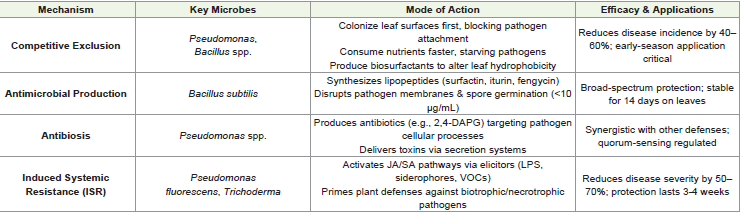
Disease Protection: The Phyllosphere as a Biological Shield:
Phyllosphere microbes protect plants through multiple
antagonistic mechanisms that offer sustainable alternatives to
chemical pesticidesCompetitive Exclusion:: Beneficial microbes establish themselves
on plant leaf surfaces before pathogens can colonize, physically
blocking their attachment. They consume available nutrients through
faster growth rates, starving potential invaders of essential resources.
Some species produce biosurfactants that alter leaf surface properties,
making them inhospitable for pathogen establishment. This spatial
dominance is particularly effective against foliar pathogens that
require specific entry points. Recent studies show that applying
competitive exclusion consortia can reduce disease incidence by 40-
60%. Field trials demonstrate that early-season microbial colonization
is crucial for maximum protective effects [64,65].
Antimicrobial Production:: Bacillus subtilis and related species
synthesize a potent arsenal of antifungal compounds including
lipopeptides like surfactin, iturin, and fengycin. These molecules
disrupt pathogen cell membranes and inhibit spore germination
at concentrations as low as 10 μg/mL. The broad-spectrum activity
of these compounds makes them effective against multiple plant
pathogens simultaneously. Formulations containing antimicrobial producing
strains maintain stability on leaf surfaces for up to 14
days post-application. Researchers are now engineering strains
with enhanced production capabilities through metabolic pathway
optimization [66,67].
Antibiosis:: Pseudomonasspecies employ sophisticated antibiotic
weapons like 2,4-DAPG that interfere with pathogen cellular
processes at multiple levels. These antibiotics work synergistically
with other microbial defense mechanisms, creating a multi-layered
protection system. The antibiotics are produced in response to
pathogen presence through quorum-sensing mechanisms, ensuring
efficient resource use. Some strains can deliver antibiotics directly to
pathogen cells through specialized secretion systems[68].
Induced Systemic Resistance (ISR):: Phyllosphere microbes trigger a sophisticated immune response in plants by stimulating
jasmonic acid (JA) and salicylic acid (SA) signalling pathways,
effectively primingthe plant’s defence mechanisms. When
beneficial bacteria like Pseudomonas fluorescens or fungi
like Trichoderma colonize leaf surfaces, they produce elicitors such
as lipopolysaccharides, siderophores, or volatile organic compounds
that activate systemic resistance. This priming effect enables plants to
respond more rapidly and strongly to pathogen attacks, with studies
showing ISR can reduce disease severity by 50-70%. The JA pathway
particularly enhances defense against necrotrophic pathogens
like Alternaria sesami, while the SA pathway targets biotrophic threats
such as powdery mildew. Importantly, ISR does not divert energy from
plant growth, making it an energy-efficient defense strategy. Field
applications of ISR-inducing microbes have shown protection lasting
3-4 weeks post-treatment. Recent advances include combining ISRinducing
strains with chitosan-based formulations that both enhance
microbial adhesion to leaves and themselves act as resistance elicitors.
Breeding programs are now selecting plant varieties with enhanced
responsiveness to ISR induction, creating synergistic plant-microbe
partnerships for sustainable disease management[69,70,71]. (Table 3)
represents key mechanisms of phyllosphere-mediated disease
protection in sesame, highlighting microbial strategies and their
efficacy. The table summarizes competitive exclusion, antimicrobial
production, antibiosis, and induced systemic resistance (ISR) as
sustainable alternatives to chemical pesticides.
Phyllosphere-Mediated Drought Resilience: Microbial Mechanisms and Applications:
formation:Phyllosphere microbes form intricate biofilm
matrices on leaf surfaces that act as natural moisture barriers.
These biofilms consist of extracellular polymeric substances (EPS)
that can hold up to 10 times their weight in water, creating a
localized humid microenvironment. The biofilm structure reduces
cuticular transpiration by 15-30%, significantly decreasing water
loss during drought periods. Certain bacterial species like Bacillus
aryabhattaiproduce hygroscopic compounds that actively capture
atmospheric moisture at night. Research shows that applying biofilmforming
consortia can improve leaf water retention by 25% under
water-deficit conditions. Farmers in arid regions are testing these
microbes as “living mulches” that can be sprayed onto crops. Recent
advances include combining biofilm formers with water-absorbing
polymers for enhanced drought protection [72,73].ACC Deaminase Activity:: ACC deaminase-producing bacteria
such as Methylobacterium and Pseudomonas play a crucial
role in drought stress mitigation by regulating plant ethylene
levels. These microbes actively cleave the ethylene precursor
1-aminocyclopropane-1-carboxylic acid (ACC), reducing stressinduced
ethylene accumulation by 40-60%. Field trials demonstrate
that ACC deaminase-containing inoculants can improve root growth
by 35% under water stress, enabling better water exploration. The
bacteria also stimulate the production of stress-responsive osmolytes
like proline and glycine betaine in plants. New formulations combine
ACC deaminase producers with mycorrhizal fungi for synergistic
drought protection. Breeding programs are now selecting plant
varieties that better recruit these beneficial microbes under stress
conditions [74,75].
UV Radiation Protection:: UV-resistant phyllosphere inhabitants like Deinococcus radiodurans and Sphingomonas species
produce protective pigments including melanins, carotenoids, and
mycosporine-like amino acids. These compounds absorb 85-95%
of harmful UV-B radiation before it can damage plant tissues. The
pigments also quench reactive oxygen species, reducing oxidative
damage to leaf cells. Some pigmented bacteria increase their UVprotective
compound production by up to 300% when exposed to
strong sunlight. Agricultural applications include foliar sprays of
pigment-producing microbes before anticipated high-UV periods.
Researchers are developing microbial consortia where pigment
producers work alongside other beneficial species for comprehensive
protection [76]
Phyllosphere microbes combat UV-induced oxidative stress
through robust antioxidant systems. SOD and catalase enzymes from
epiphytic bacteria can neutralize up to 70% of reactive oxygen species
generated during UV exposure. Some strains like Methylobacterium
extorquens increase their antioxidant enzyme production by
5-fold under high light stress. These microbial antioxidants work
synergistically with the plant’s own defense systems, providing an
additional protective layer. Field applications show that antioxidant producing
microbes can reduce UV-induced yield losses by 15-25%.
New formulations combine these microbes with natural antioxidant
compounds like flavonoids for enhanced protection [77]
Thermotolerance:Heat-adapted phyllosphere yeasts like
Rhodotorula and Cryptococcus species produce small heat-shock
proteins (sHSPs) that stabilize plant cellular structures during
temperature extremes. These microbial chaperones help maintain
the functionality of critical enzymes and membrane integrity at
temperatures up to 45°C. Some thermotolerant microbes also
induce the plant’s own heat-shock protein production through
signalling molecules. Field studies demonstrate that heat-adapted
microbial inoculants can improve pollen viability by 30% during
heat waves. Researchers are developing regional-specific microbial
blends adapted to local temperature patterns. Emerging technologies
include encapsulating these microbes in temperature-responsive
materials that release them during heat stress events [78,79].
(Table 4) depicts mechanisms of phyllosphere-mediated drought
resilience in sesame through microbial interventions. It summarizes
four key microbial strategies (biofilm formation, ACC deaminase
activity, UV protection, and thermotolerance) that enhance plant
survival under abiotic stress. Each mechanism is characterized by its
specific microorganisms, mode of action, and demonstrated efficacy
in field applications. Data are compiled from recent studies [72-79]
showing how phyllosphere microbes can be harnessed for sustainable
crop protection in water-limited environments.
Growth Promotion: Phyllosphere Microbes as Biochemical Stimulants:
The phyllosphere microbiome serves as a natural biochemical
factory, enhancing plant growth and productivity through multiple
mechanisms. These microbial stimulants offer sustainable alternatives
to synthetic growth regulators while improving crop resilience and
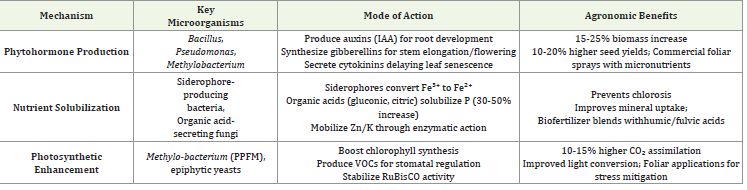
yield potential.Phytohormone Production: Microbial Growth Regulators::
Phyllosphere bacteria such as Bacillus, Pseudomonas,
and Methylobacterium play a pivotal role in development through the
synthesis of key plant hormones, each contributing distinct growth
benefits. Auxins (IAA), produced by these microbes, stimulate root
elongation, lateral branching, and vascular tissue development,
thereby enhancing the plant’s capacity for nutrient and water
uptake. Gibberellins, another class of microbial-derived hormones,
promote stem elongation, flowering, and seed set, which are critical
factors for achieving high-yield. Additionally, cytokinins secreted
by phyllosphere bacteria delay leaf senescence, ensuring prolonged
photosynthetic activity during the crucial grain-filling stage. The
practical application of these hormone-producing microbial consortia
has demonstrated significant agronomic benefits, with field trials
reporting 15–25% increases in biomass and 10–20% improvements
in seed yields. Recognizing these advantages, the agricultural
industry has begun commercializing foliar sprays that combine IAAproducing
bacterial strains with essential micronutrients, offering
farmers an effective organic alternative to synthetic growth boosters.
These microbial-based solutions not only enhance crop productivity
but also align with sustainable farming practices by reducing reliance
on chemical inputs [80,81].Nutrient Solubilization: Unlocking Mineral Availability::
Specialized phyllosphere microbes play a crucial role in nutrient
solubilization, transforming insoluble minerals into plant accessible
forms through multiple mechanisms. These beneficial
microorganisms produce siderophores - iron-chelating compounds
that convert Fe³+ into soluble Fe²+, effectively preventing chlorosis in
calcareous soils where iron deficiency commonly occurs. Additionally,
they secrete organic acids such as gluconic and citric acid that
dissolve bound phosphorus, increasing its availability by 30-50%
for plant uptake. Certain epiphytic fungi further enhance nutrient
mobilization by converting mineral-bound zinc and potassium into
bioavailable forms through enzymatic action and acidification of the
leaf surface microenvironment. Recognizing these valuable functions,
agricultural innovators have developed next-generation biofertilizer
blends that combine these nutrient-solubilizing microbes with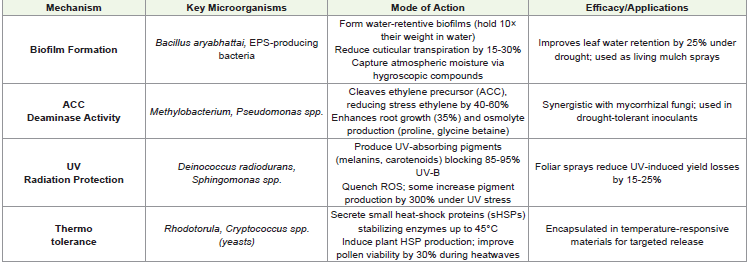
organic carriers like humic acids and fulvic acids, creating versatile
formulations suitable for both foliar application and soil treatment.
These microbial solutions not only improve plant nutrition but also
significantly reduce dependence on synthetic fertilizers, offering
a sustainable approach to crop management that maintains soil
health while optimizing productivity. The strategic use of these
microbial consortia is particularly valuable in regions with nutrient deficient
soils or where chemical fertilizer use is being curtailed for
environmental reasons [82-84].
Enhanced Photosynthetic Efficiency:The phyllosphere
microbiome significantly enhances photosynthetic efficiency through
multiple synergistic mechanisms mediated by beneficial microbes
like pink-pigmented Methylobacterium (PPFM) and other leaf associated
symbionts. These microorganisms optimize light energy
conversion by actively supporting chlorophyll synthesis, with PPFM
bacteria providing essential precursor molecules such as pyrrole
rings that boost chlorophyll production and increase leaf greenness,
typically raising SPAD values by 5-8 points. Beyond pigment support,
phyllosphere microbes improve gas exchange through the production
of volatile organic compounds like 2,3-butanediol that enhance
stomatal regulation, maintaining optimal stomatal conductance
even under environmental stress conditions. Certain epiphytic
yeasts further contribute to photosynthetic efficiency by stabilizing
and activating RuBisCO, the key enzyme in carbon fixation, thereby
increasing CO₂ assimilation rates. Field trials incorporating these
photosynthetic efficiency-enhancing microbes have demonstrated
measurable improvements of 10-15% in canopy-level carbon
assimilation, which directly correlates with higher yields. This
microbial-mediated enhancement of photosynthetic performance is
particularly valuable under suboptimal growing conditions, where
natural photosynthetic capacity may be limited, offering farmers a
biological tool to maximize the crop’s energy capture and conversion
potential without genetic modification or chemical inputs [85-87].
(Table 5) represents mechanisms of phyllosphere-mediated growth
promotion in sesame through microbial biochemical stimulants. The
table outlines three key strategies (phytohormone production, nutrient
solubilization, and photosynthetic enhancement) employed by leaf associated
microbes to boost plant productivity. Each mechanism is
characterized by its specific microbial agents, biochemical processes,
demonstrated agronomic benefits, and practical field applications.
Phyllosphere microbiota: A Sustainable Alternative to Chemical Fertilizers and Pesticides:
The phyllosphere microbiome presents a powerful, eco-friendly
solution to reduce dependence on synthetic agrochemicals in
cultivation. These leaf-associated microbes offer dual functionality,
serving as both natural biofertilizers and biopesticides. Nitrogen fixing
microbes like Azotobacter work synergistically with diseasesuppressing
Pseudomonas strains, creating multifunctional
inoculants that simultaneously enhance plant nutrition and provide
pathogen protection. This biofertilizer synergy not only improves crop
health but also reduces input costs by combining multiple benefits in
a single application. The market now offers ready-to-use biocontrol
formulations, including commercial Bacillus subtilis products that
effectively manage Cercospora leaf spot and Trichoderma-based
sprays that control Alternaria blight, all while leaving no harmful
chemical residues [88-90].These microbial solutions work through an integrated, layered
defence system that mirrors natural plant protection mechanisms.
The first line of Défense comes from competitive exclusion,
where beneficial microbes physically occupy space and consume
nutrients that would otherwise support pathogen growth. This preemptive
protection is complemented by direct suppression through
antimicrobial compounds like lipopeptides and antibiotics such as
2,4-DAPG. The system extends beyond immediate pathogen control
by inducing systemic resistance (ISR), priming the plant’s immune
system through jasmonic and salicylic acid pathways for enhanced
future protection. Together, these mechanisms-competitive
exclusion, antimicrobial production, antibiosis, and ISR-create
a comprehensive, self-reinforcing defense network. Over time, this
approach fosters long-term resilience by establishing robust microbial
communities that adapt to changing environmental conditions and
pathogen pressures.
The transition to phyllosphere-based crop protection offers
significant advantages over conventional chemical approaches.
Unlike synthetic pesticides that often lead to resistance development
in pathogens, microbial consortia employ multiple simultaneous
modes of action that are difficult for pathogens to evade. Furthermore,
these living solutions continue to proliferate and adapt on plant
surfaces, providing ongoing protection rather than the temporary
effect of chemical sprays. When combined with proper cultural
practices and monitoring, phyllosphere microbiota management
can reduce pesticide use by 40-80% while maintaining or improving
yield quality and quantity. This paradigm shift toward microbiome based
agriculture aligns with global demands for sustainable food
production, offering farmers effective tools that protect both crop
health and environmental quality.
Future Perspectives in Phyllosphere Microbiome Applications for Sesame Cultivation:
The frontier of agricultural microbiome management now
incorporates cutting-edge technologies that enable precise, data driven
microbial interventions. These innovations transform how
farmers harness phyllosphere microbes for maximum crop benefit
while optimizing resource use, particularly in developing stress resilient
sesame varieties through microbiome-assisted breeding
programs.Mechanistic Understanding of Plant-Microbe Interactions Under Field Conditions:: Future research must elucidate the precise
molecular mechanisms governing phyllosphere microbe relationships
in real-world agricultural settings. This requires long-term field
studies tracking microbial succession patterns across different growth
stages and environmental conditions. Advanced imaging techniques
like fluorescence in situ hybridization (FISH) could visualize
microbial colonization dynamics on leaf surfaces. Understanding
these interactions will enable predictive modelling of microbiome
assembly and function under various management practices. Such
knowledge is critical for developing reliable microbiome-based
solutions that perform consistently across diverse farming systems.
Standardization of Microbial Consortia for Different Agro- Climatic Zones:
The next decade will see concerted efforts to develop region specific
microbial formulations tailored to local environmental
stresses and soil types. This requires extensive field trials mapping
microbial performance across temperature, humidity, and UV
radiation gradients. Researchers must establish quality control
protocols for microbial viability during formulation, storage, and
application. Standardization efforts should include compatibility
testing with common agronomic practices in each region. Success
will depend on creating modular consortia that can be adjusted based
on real-time environmental data and crop needs.Integration of Multi-Omics Approaches for Microbial Identification:
Cutting-edge omics technologies will transform our ability
to identify and harness key functional microbes. Metagenomics
can reveal unculturable microbial taxa with beneficial traits, while
metabolomics will decode the chemical dialogue between plants
and microbes. Proteomic analyses can identify microbial enzymes
involved in stress mitigation and growth promotion. Systems
biology approaches integrating these datasets will enable the design
of synthetic microbial communities with predictable functions. This
multi-omics pipeline should become routine in microbial product
development cycles.Microbiome-Assisted Breeding Integration:
Leading seed companies are now incorporating phyllosphere
microbiome compatibility as a core selection trait in developing
new varieties.Screening thousands of genotypes for their ability
to recruit beneficial stress-alleviating microbes. Selecting for
traits that enhance microbial colonization (root exudate profiles,
leaf surface characteristics). Developing varieties that maintain
robust phyllosphere communities under drought and heat stress.
Creating customized microbial packages tailored to specific variety
characteristicsAdvanced Nano-Formulation Technologies:
Innovative encapsulation methods protect sensitive microbes
during application and ensure controlled release on leaf surfaces,
particularly important for delivering stress-alleviating microbes
in challenging conditions. Multi-layered nanocoatings respond to
environmental triggers (humidity, temperature) to time microbial
release when plants need them most. These technologies are being
adapted specifically for drought-tolerant varieties to enhance their
natural microbiome associations. Some advanced systems now
incorporate stress-specific microbial consortia with nutrient-rich
matrices that support plant-microbe symbiosis during critical growth
stages.Policy Frameworks and Farmer Adoption Strategies:
Scaling microbiome technologies requires parallel development
of supportive policies and extension services. Regulatory agencies
need science-based guidelines for evaluating microbial product safety
and efficacy claims. Governments should incentivize microbiome
technology adoption through subsidies and risk-sharing mechanisms.
Extension programs must train farmers in proper microbial product
storage, application timing, and efficacy monitoring. Demonstration
farms showcasing successful microbiome integration can build
confidence among smallholder farmers. Public-private partnerships
will be essential to make these solutions accessible and affordable
across different farm scales.Sensor Networks for Real-Time Monitoring:
Advanced hyperspectral cameras detect subtle changes in leaf
reflectance patterns that correlate with microbial activity and plant
health status. IoT-enabled smart leaf sensors continuously track
microenvironmental conditions (humidity, temperature, light)
and phyllosphere microbial dynamics, with particular attention to
stress-responsive microbial communities. These systems provide
early warnings of microbial community imbalances or plant stress
responses, allowing breeders to identify superior plant-microbe
combinations. Wireless sensor networks across fields create detailed
spatial maps of microbiome effectiveness under different stress
conditions. This real-time feedback is revolutionizing selection
processes in breeding programs focused on drought and heat
tolerance.AI-Powered Predictive Analytics:
Machine learning algorithms process historical and real-time
data on weather patterns, soil conditions, and plant phenology to
predict optimal microbial application windows. AI models analyze
complex interactions between specific varieties and their associated
phyllosphere microbiomes, identifying key microbial markers for
stress tolerance. Predictive systems now inform breeding decisions
by evaluating how different genotypes recruit and maintain
beneficial phyllosphere communities under stress. These analytics
help seed companies develop varieties with enhanced microbiome
compatibility, reducing unnecessary applications by 30-40% while
improving treatment timing accuracy for stress-prone environments.Conclusions
The phyllosphere microbiome represents an untapped reservoir
of beneficial microbes that can drive the next revolution in sustainable
agriculture. As research continues to unravel the complex interactions
between plants and their associated microbial communities, it
becomes increasingly clear that these invisible partners hold the key
to addressing some of modern agriculture’s most pressing challengesfrom
disease management and abiotic stress resilience to reducing
dependence on chemical inputs. The diverse functional roles of
phyllosphere microbes, including biocontrol, growth promotion, and
stress mitigation, offer a holistic approach to crop improvement that
works with, rather than against, natural ecosystems.
Recent advances in microbiome-assisted breeding, precision
application technologies, and microbial consortia development are
rapidly translating laboratory discoveries into practical farming
solutions. The integration of multi-omics approaches with traditional
agricultural knowledge is creating new opportunities to customize
microbial interventions for specific varieties, growth stages, and
environmental conditions. However, realizing the full potential
of phyllosphere microbiome engineering will require overcoming
significant challenges in standardization, scalability, and farmer
adoption.
As we move forward, the successful implementation of
microbiome-based agriculture in sesame production will depend on
three critical factors: (1) continued research into the fundamental
ecology of plant-microbe interactions under field conditions, (2)
development of robust, climate-smart microbial formulations
that maintain efficacy across diverse growing regions, and (3)
establishment of supportive policy frameworks that facilitate
technology transfer to farmers. The interdisciplinary nature of this
work-bridging microbiology, plant science, data analytics, and
social sciences-underscores both its complexity and its tremendous
potential.Ultimately, harnessing the phyllo sphere microbiome
represents more than just a novel agricultural strategy-it embodies
a paradigm shift toward working with nature’s own systems to
create more resilient, productive, and sustainable food production
systems. As climate change intensifies and global demand for sesame
continues to grow, these microbial solutions may prove indispensable
for ensuring food security while protecting environmental health.
References
Citation
Prasanna Lakshmi B, Sreeramulu A. The Plant Microbiome: Harnessing Phyllosphere Microbial Communities for Sustainable Cultivation of Sesame (Sesamum indicum). J Plant Sci Res. 2025;12(2): 280