Research Article
Agrobacterium Mediated Hairy Root Induction in Lawsonia inermis L.: A Step Forward for Secondary Metabolites Production
Moharana A1,2*
1Department of Botany, Ravenshaw University, Cuttack-753003, Odisha, India.
2ICAR-National Rice Research Institute (ICAR-NRRI), Cuttack-753006, Odisha, India
2ICAR-National Rice Research Institute (ICAR-NRRI), Cuttack-753006, Odisha, India
*Corresponding author:Arpita Moharana, ICAR-National Rice Research Institute (ICAR-NRRI), Cuttack-753006, Odisha, India. E-mail Id: arpiarpita22k@gmail.com
Copyright: © Moharana A. 2025. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Article Information: Submission: 03/01/2025; Accepted: 05/02/2025; Published: 07/02/2025
Abstract
In this study, a protocol for hairy root production for bioactive compound enhancement of Lawsonia inermis through intervention of Agrobacterium rhizogenes was attempted. Two types of explants, leaf (both in vivo and in vitro) and internode (both in vivo and in vitro) were taken for hairy root transformation
by two Agrobacterium rhizogenes strains (MTCC 532 and MTCC 2364). Among leaf and internode, better transformation observed in leaf than internode where as in vitro leaf was found most suitable explant for optimum transformation than in vivo leaf explant. Better transformation was achieved by MTCC
2364 than MTCC 532 in all leaf and internodal explants. Highest 83.3% hairy root induction was noticed in in-vitro leaves infected by MTCC 2364 for 40 minutes infection time with an O.D. value 0.6 and co-cultivation time 24-26 hours. Root emergence in in vitro leaf started within 15-17 days and highest c.a.9.2 roots were observed. The in vivo leaf was found to develop hairy roots within 20-23 days from the day of infection with 73.3% transformation efficiency infected by MTCC 2364 and developed c.a 8.4 roots.
Keywords:Agrobacterium rhizogenes strains (MTCC 532 and MTCC2364); In vivo leaf and internode; In vitro leaf and internode; Secondary metabolites
Introduction
Lawsonia inermis L. (syn. Lawsonia alba Lam. or Lawsoniaruba
L.), commonly known as Henna or Mehendi [1,2] is a monotypic
genus of family Lythraceae. India’s ancient history discusses its
many applications and significant importance in Ayurvedic or natural
herbal remedies. Apart from cosmetic use, Mehendi leaves are used
as a prophylactic against diarrhea, skin diseases, renal lithiase, and
gastric problems. The flowers have also medicinal properties, used as
refrigerant, soporific, febrifuge, cardio tonic, as an emmenagogue and
applied against bruises. The seed powder is effective for dysentery,
liver disorders, and associated problems. Besides the aerial plant
part, the root plays an immensely significant role in the treatment
of various diseases. Root has astringent properties and is used for
sore eyes, bruises, and boils of children’s heads. The root of this
plant is helpful in the treatment of hysteria, gonorrhea, herpes,
tumor, nervous disorders, and improves the liver and kidney [3-5].
Most importantly, in Odisha and in particular Koraput district and
Kandhamal district, the root has also been used for the treatment of
jaundice by common people, tribal people as well as local traditional
healers [6,7]. They used root paste of L. inermis along with raw
rice water for treatment of jaundice. Besides, pharmacologically
the methanol extract of roots of L. inermis was most effective as
an abortant [3] ethanolic root extract is effective as an antitumor,
antiproliferative and has been used to improve the hepatic and renal
function.[8,9] In addition, the presence of important secondary
metabolites in the aerial part (in vivo and in vitro) as well as in root (in
vivo and in vitro) has been proven by High Performance Thin Layer
Chromatography (HPTLC; Moharana et al., 2018b) [10]. Due to the
immense significant traditional and pharmacological properties of
L. inermis root, its popularization as herbal medicine and extraction
of secondary metabolites for pharmacological uses is necessary.
For pharmaceutical commercialization, large scale production of L.
inermis root is a prerequisite.
In this context, in vitro root cultures in particular hairy root
production could be the most appropriate alternative method.
Biotechnological intervention through Agrobacterium rhizogenes can
be used as a strategy for a sustainable industrial scale root production.
A. rhizogenes (now known as Rhizobium rhizogenes), a soil-dwelling
Gram-negative phytopathogenic bacteria when infect the plant
caused “hairy root” disease. As a consequence, uncontrolled root
growth at the site of injury and infection of the plant is found. The
additional advantages of the production of secondary metabolites
through in vitro hairy root culture includes fast growth, low doubling
time, ease of maintenance of hairy roots, and genetic stability [11].
The biosynthetic ability of “hairy root culture” to produce secondary
metabolites is often equal to or greater than mother plant. [12-14]
Interestingly, hairy root has also been proven to synthesize those
compounds which are known to accumulate in aerial part only.
[15,16,14]
In short, L. innermis is a traditional medicinal plant of which root
is a most important part attributing many pharmacological activities.
Therefore, an increase in root biomass without uprooting the plant is a
prerequisite for industrial-scale production of secondary metabolites
particularly located in the root. So, in this part of work attempt has
been taken to increase root biomass through hairy root cultures of
L. inermis using A. rhizogenes for production and enhancement of
secondary metabolites located in the root. Apart from that, factors
influencing genetic transformation including explant source and
type, culture matrix, bacterial strains, bacterial cell density, method of
infection, and co-cultivation period were evaluated and standardized
to maximize the efficiency of the transformation and hairy root
production in L. S.
Material and Methods
Explant source (both in vivo and in vitro) for Agrobacterium transformation:
For Agrobacterium infection experiment, two types of explants
i.e., leaf and internode (mature and axenic) were taken. Young, fresh
leaves and internodes were collected from a six years old L. inermis
plant (Figure1A) present in campus of Ravenshaw University. The
plant specimen which is used as the mature source (in vivo) of
explants was identified and deposited in the herbarium of Botanical
Survey, Odisha (voucher specimen number 2539/CBT). The mature
leaves and internodal explants were kept under running tap water
separately in a beaker with constant shaking of about 30 min for
removal of external adherents like dust and dirt. Then, both types of
explants were washed for 5 min with 1 % (v/v) aqueous solution of
Teepol (Reckilt Benckiser Ltd., India), followed by five times rinsing
with double distilled water. Then a 0.1 % (w/v) aqueous solution of
mercuric chloride (HgCl2, Hi-Media, India) was used for surface
sterilization (3 min for leaves and 4 min for internodes), followed
by five-six rinsing of sterile double distilled water [5]. After rinse,
both leaves and internodal explants were ready for Agrobacterium
infection.For axenic (in vitro) internode and leaf, the in vitro regenerated
plantlets maintained in our laboratory (Figure 1B) were used as a source
of explant. The in vitro shoot culture was established by inoculation of

in vivo nodal explant on Murashige and Skoog, medium (MS, 1962)
supplemented with BA (1.0mg/ml) and subsequent elongation on MS
medium devoid of any plant growth regulators (PGRs) [5].
Bacterial strains and their maintenance:
Two types of strains of Agrobacterium rhizogenes, MTCC 532 and
MTCC 2364, were procured from IMTECH (Institute of Microbial
Technology), Chandigarh, India and revived in nutrient broth (Hi-
Media). For bacterial growth, the temperature was set at 26±1°C for
44 to 48 hours inside the incubator. The broth cultures were stored at
4 °C for the Agrobacterium transformation experiment. For explant
infection, a 100μl aliquot of bacterial suspension was added into 50
ml of liquid medium in a 150-ml Erlenmeyer flask and incubated in
an incubator shaker at 26±1°C. Prior to bacterial inoculation, the OD
of both the bacterial suspensions of both the strains was adjusted to
0.6 (at 660 nm).Agrobacterium transformation and hairy root induction in leaves (in vivo and in vitro) and internodes (in vivo and in vitro):
For the transformation and hairy root induction experiment,
thein vivo internodal segments were excised (1.5-2.0cm) from both
sides to eliminate the damaged tissue that were affected during the
process of surface sterilization. The axenic internodes were excised
(1.5-2.0cm) from in vitro regenerated shoots. Then both types of
internodes were punctured on one tip side end, followed by the
dipping in the bacterial suspension for 10-60 minutes to allow the
infection. Likewise, leaves were excised from in vitro shoot culture,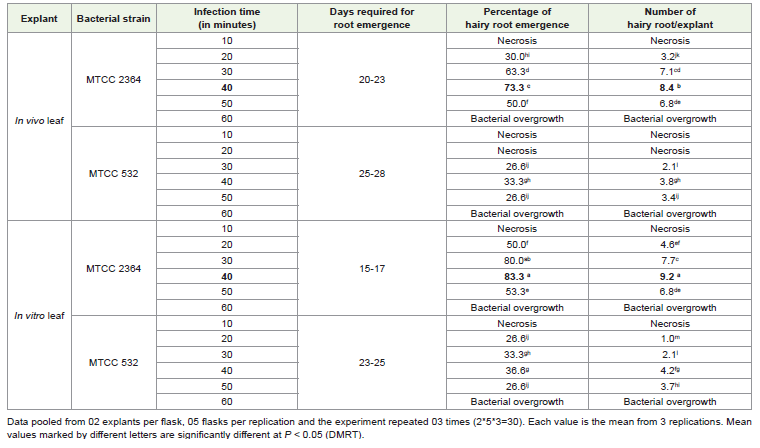
punctured around the midrib and dipped in the bacterial suspension
for 10-60 minutes for Agrobacterium infection.
Both leaves (in vivo and in vitro) and internodes (in vivo and
in vitro) were taken out of the bacterial suspension and rinsed with
sterile distilled water, followed by drying with sterile tissue paper.
Internodes were inoculated by the other side of the piercing and
leaves were by their dorsal side up on solid half MS (agar 0.7 %, Hi-
Media; pH 5.8±0.01). The co-cultivation time was optimized between
24-26 hours inside an incubator at 26±1°C. After co-cultivation,
explants were transferred to fresh flask with solid half MS (agar 0.7 %,
pH 5.8±0.01) and kept inside a dark culture room at 25±1°C for hairy
root induction. One set of controls was maintained by the inoculation
of both types of leaves and internode separately on solid half MS
medium without exposure to bacterial suspension.
Establishment and mass enhancement of hairy root:
After hairy root induction from both in vivo leaf and in vitro
leaf, individual transformed roots (1.5–2.0 cm) were excised and
transferred to a conical flask (Borosil, India) containing 100ml of
liquid ½ MS medium without antibiotics incubated inside a dark
culture room at 25±1°C. At one week interval, the hairy roots were
transferred into a freshly prepared liquid ½ MS medium without
antibiotics in the same conditions for root biomass enhancement.Experimental design, statistical analysis and photography:
For hairy root development, each treatment of the experiment,
consisted of 5 replicates (culture flasks) and two explants/flask was
the experimental unit. Each experiment was repeated thrice at a 5 days
interval. Mean value was taken from three biological replications.
The percentage of explant showing hairy root induction, number of
roots/explants, and root length were recorded after 45 days by visual
observations. Mean values within column with different superscript
alphabets are significantly different. Data were analyzed by analysis
of variance (ANOVA) using Duncan’s multiple range test (p < 0.05).
Explants with hairy roots were photographed by a Canon DSLR P3000
camera and uploaded to the computer by inserting the memory card
into computer.Results
Agrobacterium transformation and hairy root induction in leaves (in vivo and in vitro)::
For Agrobacterium transformation, both mature and axenic
leaves, were taken into consideration. No hairy root development was
observed in the control set of the experiment. Early rooting response
and higher root regeneration frequency were observed in in vitro
leaves than in vivo leaves [Table 1]. The hairy root development in
both types of leaves was observed mostly from the cut ends of the
petiole and midrib. Root induction occurred in 15-17 days in in vitro
leaf whereas, 20–23 days were required for rhizogenesis of in vivo leaf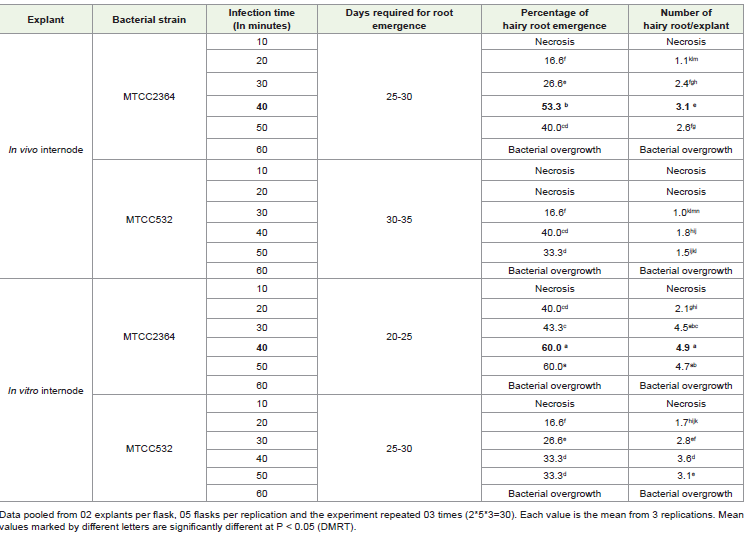
[Table 1] infected by MTCC 2364. The in vitro leaves resulted in the
highest 8.5 (Figure 1C) hairy roots whereas, mature leaf developed
the highest 7.2 (Figure 1D) hairy roots. Bacterial contamination was
found more in in vivo explants, where as in vitro explants was devoid
of such types of contamination.
Among the two MTCC 532 and MTCC 2364 A. rhizogenes strains
used for hairy root formation, MTCC 2364 found much better in
hairy root induction irrespective of explant types. The microbial
strain MTCC 532, resulted in a lower percentage of root regeneration
with few numbers of roots whereas, MTCC 2364 showed a higher
transformation frequency in both in vivo (73.3) and in vitro (83.3)
leaf. Apart from that, a comparatively higher number of days are
required for rhizogenesis in the case of MTCC 532 than MTCC 2364
irrespective of explant types [Table 1].
Out of different infection time periods, 40 min. was found
to be more effective in terms of the highest number of hairy root
regeneration both in in vivo (8.4) and in vitro (9.2) leaves, whereas
much lower infection time (10 mins.) did not show any response in
any type of leaf explant rather the leaves eventually became brown
and died after a few days of infection. Leaf explants infected for more
time (60 min.) found excess bacterial growth around them with no
hairy root [Table 1].
The O.D. values for both the strains were optimized (data not
shown) to 0.6 at 660 nm and co-cultivation time as 24-26 hours (data
not shown) for optimum transformation in both types of leaves.
Agrobacterium transformation and hairy root induction in internodes (in vivo and in vitro):
Out of in vivo and in vitro internodes, higher transformation
by Agrobacterium was observed in in vitro internodes than in vivo
internodes. The in vitro internodal explant showed an early rooting
response than the in vivo explant. The hairy root development in
both types of internodes was observed mostly from the piercing cut
end. Root induction occurred in 20-25 days on in vitro internodes
whereas, 25–30 days were required for root emergence on internodes
from mature explants (in vivo) (Table 2) infected by MTCC 2364. The
in vitro and in vivo internodes infected by MTCC 532 showed root
emergence in 25-30 and 30-35 respectively. The in vitro internodes
resulted in the highest 3.1 (Figure 1E) hairy roots whereas, mature
internodes developed the highest 4.8 (Figure.1F) hairy roots.
Both types of internodal explants were found devoid of bacterial
contamination. No hairy root development was observed in the
control set of the experiment.Among MTCC 532 and MTCC 2364 A. rhizogenes strains used
for hairy root induction, MTCC 2364 found much better in hairy root
development irrespective of explant types. The microbial strain MTCC
532, resulted in a lower percentage of root regeneration with a few
numbers of roots whereas MTCC 2364 showed higher transformation
frequency in both in vivo and in vitro internodes. Apart from that, a
greater number of days are required for rhizogenesis in the case of
MTCC 532 than MTCC 2364 irrespective of explant types [Table 2].
Out of different infection time periods, 40 min. was found to be
more effective in terms of highest number of hairy root regenerations
both in in vivo (3.1) and in vitro (4.9) internodes whereas much lower
infection time (10 min.) did not show any response in any type of
internodal explants rather, the internodes became brown and died
after a few days of infection. The higher percentage of transformation
frequency was also recorded with 40 min. of infection time in both
in vivo (53.3 %) and in vitro (60 %) internodal explants. Internodal
explants infected for more time (60 min.) found excess bacterial
growth around them with no hairy root [Table 2].
The O.D. values for both the strains were optimized (data not
shown) to 0.6 at 660 nm and co-cultivation time as 24-26 hours (data
not shown) for optimum transformation in both types of internodes.
Establishment and mass enhancement of hairy root:
Optimum hairy root induction was resulted by the infection of
MTCC 2364 in leaf explant in terms of percentage of explant response
(73.3 % in in vivo and 83.3 %in vitro) and number (8.4 in in vivo and
9.2 in in vitro) of hairy root regeneration as a result of transformation.
So, the further experiment was carried out by taking hairy roots
developed from both types of leaves. Actively growing individual
roots (hairy roots; 1.5-2.0 cm) from transformed leaves, were excised
and transferred to liquid ½MS medium. After 2 weeks, the roots in
liquid medium showed a slight elongation as well as regeneration of
secondary roots from the primary root (Figure 1 G),(Figure 1H)Discussion
Out of the two strains, MTCC 532 and MTCC 2364 were selected
for the hairy root induction experiment, the MTCC 2364 strain of
A. rhizogenes was found more effective for hairy root induction in
comparison to MTCC 532. These two strains were also previously
used by Brijwal and Tamta, 2015[17] (Berberis aristate),Bathojuet
al., 2017[18] (Chlorophytum borivilianum) for hairy root induction.
However, out of these two strains, MTCC 532 was reported by them
as more effective in terms of percentage of hairy root induction,
biomass, number, and length of hairy root than MTCC 2364, which
is contrary to the result of this present study. At the same time,
Vishwakarma et al., 2017[14] and Mahakuret al., 2024 [19] reported
MTCC 2364 is more effective in hairy root formation than MTCC 532
in Mucuna pruriensand Vitex negundorespectively, which is similar to
the result of the present work. On the contrary, Deore and Kide, 2015
[20] (Chlorophytum species) found MTCC 2364 as totally ineffective
for hairy root development. Except for these two bacterial strains,
Bakkali et al. [15] (1997) successfully developed hairy root from the
in vitro leaf of L. innermis by the transformation of Agrobacterium
rhizogenes NCIB 8196.
The types of explants influenced the hairy root production so, with
a view to determie the suitable explant for optimum Agrobacterium
transformation, different types of explants i.e., leaf (in vivo and in
vitro) and internode (in vivo and in vitro) were used, out of which,
both in vivo and in vitro leaf explants were more responsive than both
types of internodal explants. Whereas, in vitro leaf showed better
transformation percentage as well as more number root development
than the in vivo leaf. Contrary to this result, internodal explant of
Clitoriaternatea reported as better for Agrobacterium transformation
than the leaf [21] (Swain et al., 2012). Most of the researchers,
including Thilipet al., 2015[22] (Withaniasomnifera) and Jesudasset
al., 2020[23] (Cucumis anguria) suggested the use of in vitro or
axenic explant for hairy root induction, which is similar to this part
of the experiment. Preference of in vitro explants may be to avoid
the crucial step of surface sterilization which is time consuming and
has a chance of microbial contamination. But Swain et al., 2012 [21]
(Clitoriaternatea), Srinivasan et al., 2023[23](Aerva javanica) also got
success in regeneration of transformed roots from in vivo explants.
Agrobacterium cell density is a fundamental factor influencing
genetic transformation system (Kumar et al., 2006; Binka et al., 2012;
Shahabzadeh et al., 2014; Asande et al., 2020). [24-27] In this part of
the experiment, the suitable O.D. value of the MTCC 2364 was found
to 0.6 for the hairy root induction in all types of explants. Swain et
al., 2012[21] (Clitoriaternatea) suggested a 0.6 O.D. value as suitable
for optimum hairy root production for A. rhizogenesA4T which
corroborates the result of this part of the research work. But it is a fact
that for perfect transformation the optical densities of Agrobacterium
suspension cultures ranged from 0.1 to 1.0 depends on the genotype,
Agrobacterium strain, and plant species (Asandeet al., 2020).[27]
Higher Agrobacterium density can cause uncontrolled growth of
bacterial cells thus limiting the explant survivility and subsequent
reduction in transformation efficiency. Like-wise, lower cell density
cannot induce transformation.
The transfer of T-DNA to the plant genome from Agrobacterium
during transformation process is time-dependent and therefore,
transformation efficiency depends on both time of infection and cocultivation
duration (Markandan et al., 2015; Asandeet al., 2020).
[27,28]Apart from that, A. rhizogenes strains are different in their
virulence, which leads to the different development rates of hairy
root (Giri et al., 2001).[29]Variation in the infection durations had an
influence on the transfer of T-DNA from Agrobacterium to plant cells
of L. inermis from which, 40-min of infection time was recommended
for successful transformation. Furthermore, the hairy root induction
with 20- and 30-min infection periods resulted in significantly lower
infection rates and number of roots. Due to inadequate bacterial
infection duration, a shorter infection period like 10 min. was found
ineffective. A higher rate of infectivity was not found when the
infection duration crossed 40 min. Rather, 60 min infection time
showed overgrowth of bacteria leading to explant contamination
followed by necrosis. Therefore, 40 min was determined to be the
optimal time for infection for L. inermis. Similar type of observation
was reported by Srinivasan et al. (2023) where, he suggested 20
min. as the optimum infection period for hairy root formation, but
lower than 20 min. and higher than 20 min. are not preferable. Effect
of infection time on transformation frequency using A. rhizogenes
also proven to be dependent on plant species. Five minutes of
infection to wounded explants was effective in inducing hairy roots
in Linum mucronatum (Samadi et al., 2012)[30]and in Agastache
foeniculum (Nouroziet al., 2016),[31]whereas in Silybum marianum
(Rahnama et al., 2008)[32]and in Fagopyrum tataricum (Thweet
al., 2016),[33-37]10 min of infection time was found effective for
optimum transformation. Adding to that, 20 min. in Artemisia annua
(Giri et al., 2001)[29]and one hour in Berberis aristata(Brijwal and
Tamta, 2015)[17] were required for maximum hairy root formation.
Conclusion
To give a step forward for production and enhancement of root
specific important phytochemicals, induction and establishment of
hairy root in L. inermis by by Agrobacterium mediated transformation
was carried out in which, in vitro leaf was found as the most suitable
explant for hairy root induction by A. rhizogenes MTCC 2364 with
O.D 0.6 and infection time 40 min. with co-cultivation time 24-26
hours. Factors impacting genetic transformation including explant
source and type, culture matrix, bacterial strains, bacterial cell density,
method of infection, and co-cultivation period were evaluated and
standardised to maximize the efficiency of the transformation and
hairy root production in L. inermis.
However, this part of the research work remains as a preliminary
part of the work. Further research is under process i.e., enhancement
of transformation efficiency, root number, and length. Molecular
validation of transformed roots by PCR amplification, hairy root
biomass enhancement, parameter optimization for production and
enhancement of important secondary metabolites and phytochemical
validation of secondary metabolites in both transformed root and
roots from the mother plant will be accomplish. This part of the
work might be a path forward for optimization of industrial scale
production of roots biomass or bioreactor design aiming in the
production of secondary metabolites, particularly accumulated in
root, for drug manipulation to combat the antibiotic-resistance
human pathogens, as L. inermis holds the status of a multipurpose
medicinal plant.
Declaration of Conflicting Interests and Ethics: The author
declares no conflict of interest
Funding support: The author declares that she has not taken any funding support to
carry out this part of research work.
Authorship contribution statement: The Author designed, carried out the whole experiment, analyzed
the data and wrote the entire manuscript
Acknowledgement
The author is grateful to Department of Botany, Ravenshaw
University, Cuttack-753003 for providing necessary laboratory
facilities. AM is highly obliged to Dr. Durga Prasad Barik and Dr.
Soumendra Kumar Naik for their support, guidance, and providing
the bacterial strains in carry out the experiment.
References
Citation
Moharana A. Agrobacterium Mediated Hairy Root Induction in Lawsonia inermis L.: A Step Forward for Secondary Metabolites Production. J Plant Sci Res. 2025;12(1): 273