In vitro Regeneration and Effect of Abiotic Stress on Physiology and Biochemical Content of Stevia Rebaudiana ‘Bertoni’
Mukeshwar Pandey* and Surendra K. Chikara
Corresponding author: Mukeshwar Pandey, Shri Venkateshwara University, NH-24, Vankateshwara Nagar, Rajabpur,Gajraula 244236, India, Tel: +91-8477956666,; E-mail: mukesh02.max@gmail.com
Citation: Pandey M, Chikara SK. In vitro Regeneration and Effect of Abiotic Stress on Physiology and Biochemical Content of Stevia Rebaudiana‘Bertoni’. J Plant Sci Res. 2014;1(3): 113.
Copyright © 2014 Pandey M et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Plant Science & Research | Volume: 1, Issue: 3
Submission: 03/11/2014; Accepted: 20/11/2014; Published: 21/11/2014
Abstract
The present work aim to regenerate S. rebaudiana ‘Bertoni’ under in vitro condition and also investigating the effect of salinity anddrought stress on growth, physiological response and biochemical content. Cultures were established on Murashige and Skoog (MS)medium supplemented with different concentrations of 6-benzylaminopurine (BAP) and Kinetin (Kn) individually and/or in combination.The highest frequency of bud break (90%) as well as bud elongation (95%) was obtained on MS medium supplemented with 0.5 mg/lBAP and highest mean number of shoot (30) and shoot length (12.2cm) with 1.0 mg/l BAP + 0.5 mg/l Kn along with rooting at 2.0 mg/lIBA. Moreover, when plants were subjected to NaCl and mannitol, significant decrease in shoot number, shoot length, root number androot length were observed with the increasing concentration of NaCl and mannitol (25, 50, 75 and 100mM) while significant increase wasfound in all parameters with increasing number of days (10, 20 and 30) as compared to control. In addition, shoot/root ratio, leaf no., leafdry weight, leaf fresh weight, stem dry weight, root dry weight and shoot dry weight were continuously decreased (p < 0.05) as NaCl andmannitol concentration increased. The rate of proline accumulation was continuously increased and protein content was continuouslydecreased at different concentration of NaCl and mannitol. Moreover, significant reduction in chorophyll content was also observed withincreasing concentrations of NaCl and mannitol. The results showed that salinity and drought stress significantly reduces the growth andyield components of S. rebaudiana ‘Bertoni’ by affecting endogenous growth hormones.
Keywords: In vitro regeneration; Salinity stress; Drought stress; S. Rebaudiana; Biochemical content
Abbreviations: : BAP: 6-Benzylaminopurine; NAA:α-Naphthaleneacetic acid; IAA: Indole-3 acetic acid; 2 4-D: 2,4-Dichlorophenoxyaceticacid; Kn: Kinetin; FBB: Frequency of bud break;FBE: Frequency of bud elongation; MNS: Mean no. of shoot; MSL:Mean shoot length; MNR: Mean no. of root; MRL: Mean root length
Introduction
In view of current situation of food insecurity, particularly indeveloping countries, a number of factors cause a decrease in cropproductivity. Of them, availability of agricultural land, fresh water resources, ever-increasing abiotic stresses, and low economic activityin agricultural sector are the most important factors. However, it isgenerally believed that abiotic stresses are considered to be the mainsource of yield reduction. Both water and salt stress affected more than10% of arable land, which results in rapid increase in desertificationand salinization world-wide. As a consequence, average yields ofmajor crops reduced by more than 50% [1]. Due to this reason, thereis an increasing demand for new plant cultivars that have a potentialfor higher yield under such abiotic adversaries.
Stevia [Stevia rebuaidana (Bertoni); family: Asteraceae] is a perennial shrub widely planted in many countries such as China,Japan, Korea, India and certain countries of South America [2]. It is anatural sweetener plant known as sweet wee, sweet leaf, sweet herb onhoney leaf, which is estimated to be 300 times sweeter than sugarcane[3]. Stevioside and rebaudioside are the active principal constituentsof diterpene glycosides with varying sugar molecules present inleaves of stevia plant are responsible for high sweetness. Steviosideis abundantly available in leaves (13-20% in dry leaves) with 250-350times sweetness. Rebaudioside (1-3% in dry leaves) is 350-450 timessweeter while that of fresh healthy leaves are only 30 times sweeterthan table sugar. This is used as a substitute for saccharose, and inthe treatment of diabetes mellitus, obesity, and hypertension and incaries prevention [4].
The seed of Stevia show very low germination percentage [5] andpropagation by seeds does not allow the production of homogenous population resulting in variability in sweetener level and composition. Vegetative propagation by stem cuttings is also limited by the lownumber of individuals that can be obtained simultaneously from single plant. Micropropagation can provide genetically uniform plantsin large numbers. However, with climate change and land clearing orirrigation, a large quantity of salt and alkali land have occurred inthese areas, which possessed excellent light conditions for the growthand glycoside accumulation of S. rebaudiana. Previous studieson plants under salt stress had revealed alterations in the biomass,antioxidant enzyme activities and osmolytes, mineral contents, andsecondary metabolites [6]. However, salt effects on the survival andproductivity of S. rebaudiana have rarely been reported [7]. Thus,investigating these responses is difficult under field conditions, butplant tissue culture techniques are performed under aseptic andcontrolled environmental conditions. The advantages of plant tissueculture allow various opportunities for researcher to study the uniqueand complex responses of plants against environmental stresses.
Materials and Methods
Plant Material and In vitro Regeneration
A plant of family Asteraceae was selected as experimentalmaterial, obtained in the form of Stevia rebaudiana Bertoni plantgrowing in the Botanical Garden of Anand Agricultural University,Anand (Gujarat) INDIA. Axillary bud from the actively growing newshoots were excised and used as explant. The explants were rinsed inrunning tap water for 20 min, cleaned in 10% liquid soap solutionfor 5 min and then rinsed with double distilled water 2-3 times.The explants were surface sterilized with 0.1% mercuric chloridesolution and for 2-3 min in 70% ethanol. Finally, the explants werewashed in sterilized double distilled water and cultured on MSmedium (Murashige and Skoog 1962) with vitamins, 3% sucrose,0.8% agar supplemented with different concentrations (0.5-2.0 mg/l)of 6-benzylaminopurine (BAP) and Kinetin (Kn) for establishmentand multiplication of culture. The pH of the nutrient medium wasadjusted to 5.8 and 0.8% agar (Himedia, Mumbai) was added prior toautoclaving. The regenerated shoots were cultured on half-strengthMS medium fortified with auxins viz. indole acetic acid (IAA) atdifferent concentrations (0.5-2.0 mg/l) to study their effects onrooting. All the cultures were incubated at 27±2 °C under 2,000-2,500 lux for 16 h/day.
Plantlets with well developed roots were washed thoroughly withdistilled water and cultured in a mixture of sand and peat (1:1) in potsand transferred to green house. The potted plants were irrigated andinitially covered with plastic bags, which was gradually eliminatedwithin four weeks for completing their acclimatization.
In vitro salinity and drought stress studies
In vitro salinity and drought stress conditions was developed asdescribed by Debnath et al. [8] with modification in the concentrationof growth regulators, NaCl and mannitol. For this, cluster of multipleshoots were separated and re-inoculated on MS medium supplementedwith 1 mg/l BAP + 0.5mg/l Kn and different concentration of NaCl(0, 25, 50, 75 and 100 mM) and mannitol (0, 25, 50, 75 and 100 mM)for salinity and drought stress, respectively. These growing explantswere retrieved from the modified MS medium at 10, 20 and 30 daysinterval for evaluation of plant development along with biochemicalquantification of proline, protein and chlorophyll.
Biochemical constituent’s analysis
Extraction and estimation of Proline Assessments of prolinecontent were performed twice during the experimental period, at10, 20 and 30 days after the onset of the experiment. Proline wasextracted from sample of 0.5g fresh leaf material samples in 3% (w/v)aqueous sulphosalycylic acid and estimated using the ninhydrinreagent according to the method of Bates et al. [9]. The absorbanceof fraction with toluene aspired from liquid phase was read at a wavelength of 520 nm.
Extraction and estimation of Protein Protein extraction wasperformed as described by Hurkman and Tanaka [10]. 1 g freshtissue was grinded in a chilled mortar and pestle with 2.5 mL ofTris (pH 8.8) buffered phenol and 2.5 mL of extraction media (0.1M Tris-HCl pH 8.8, 10 mM EDTA, 0.4% 2- mercaptoethanol, 0.9M sucrose). The homogenate was centrifuged for 10 min at 5000 gfor 4 °C. The aqueous phase at the bottom was mixed with 2.5 mLof extraction media and phenol by vortexing. The phenol phase atthe top of the extract was separated twice by this method and pooledtogether. Proteins was precipitated from the phenol extract by adding5 volumes of 0.1 M ammonium acetate in 100% methanol (stored at-20 °C) and centrifuging at 20,000 g, 4°C for 20 min. The precipitateprotein was incubated overnight at – 20 °C.
Protein quantification was performed as described by Lowry etal. [11]. To 1 ml of above solution, 5 ml of freshly prepared alkalineCuSO4 reagent [1 ml of Alkaline Na2CO3 reagent to 50 ml of CopperSulphate reagent] was added mixed properly and after 10 min Folin’sreagent (0.5 ml) was added. The solution was mixed instantaneouslyand allowed to develop the colour. Absorbance was recorded at 750nm.
Extraction and estimation of chlorophyll Following extraction of liquid-nitrogen frozen leaf with 80% acetone, the concentration ofchlorophyll was quantified by spectrophotometer method of Porra et al. [12].
Statistical analysis and growth evaluation
The morphogenetic responses of the explants was assessed and recorded regularly up to a maximum of 15 passages. For each treatment, 12 replicates were maintained and each experiment was repeated at least three times. The results were noted in the form ofMean ± Standard Error (SE). The growth parameters was calculated as frequency of bud break (FBB), frequency of bud elongation (FBE), mean number of shoot (MNS), mean shoot length (MSL), mean number of root (MNL) and mean root length (MRL). The data recorded were analyzed statistically using analysis of variance technique including one-way and two-way ANOVA.
Results and Discussions
In vitro Regeneration of Stevia rebaudiana ‘Bertoni’
Initiation or establishment of culture In vitro propagation studiesshow that plant hormones play an important role in regenerationof culture. For establishment of culture, different cytokinin (BAPand Kn) was used. The axillary bud explants were cultured on MSmedia supplemented with different concentration (0.5, 1.0, 1.5 and2.0 mg/l) of BAP and Kn. BAP at 0.5 mg/l stimulated proliferationof the bud meristems to form bud clusters and maximum frequencyof bud break (FBB) reached 90% and bud elongation reached 95%as compared to higher concentrations of BAP where slow growthwas observed. Similar result was obtained by Abd-Alhady [13]. Thestimulating effect of BAP on bud break has been reported earlier by Debnath [14]. In the initiation stage, Kn also showed maximumfrequencies of bud break (85%) and bud elongation (92%) at 0.5 mg/lKn (Figure 1a; Figure 2a) but lower than BAP while at concentrationgreater than 0.5 mg/l treatment resulted decline in all parameters.
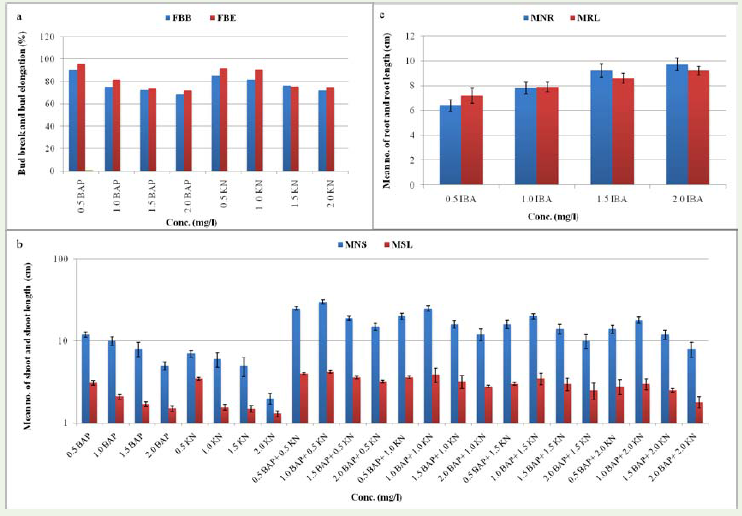
Figure 1: In vitro regeneration of Stevia rebaudiana Bertoni under aseptic medium a) Initiation or establishment of culture at different concentration (0.5, 1.0, 1.5 and 2.0mg/l) of BAP and Kn b) Multiplication of explants at different concentration and combination of BAP and Kn c) Rooting of plantlets at different concentrationof IBA (0.5, 1.0, 1.5 and 2.0mg/l).
Multiplication and elongation of Shoots The proliferation and multiplication of the shoots were also influenced by hormones whenadded singly or in combination. The mean number of shoot (MNS) per explant and mean shoot length (MSL) varied under various cytokinin concentrations and combinations (Figure 1b; Figure 2b, c). The effect of different concentration of BAP used was found to be significantfor increasing the number of shoots (mean number of shoot, MNS) after 3-4 weeks of inoculation. Maximum number of shoots (14±0.94) and shoot length (10.2±0.83) was observed at 0.5mg/l concentration while at concentration higher than 0.5 mg/l treatment, the shoot number declined. It was clear that among different concentration of Kn used, only at 0.5 mg/l Kn concentration, the highest numberof shoots (12±0.65) and shoot length (10.4±0.14) after 3-4 weeks was observed while at higher concentration than 0.5 mg/l Kn, the shoots number declined. Between the two cytokinins tested, BAP was more effective than Kn for shoot multiplication. When cultures were subjected to different combinations and concentrations of BAPand Kn, significant changes were observed. Our data revealed that Stevia rebaudiana, Bertoni cultures treated with 1.0 mg/l BAP in combination with 0.5 mg/l Kn found significant increase in shoot number (30±1.83) and shoot length (12.2±1.83) while concentrationlower or higher declined MNS and MSL (figure 1b). Abd-Alhady [13] reported highest shoot multiplication in medium supplementedwith 1.0 mg/l BAP + 0.5 mg/l Kn (36.9 shoots per explant) and lowest shoot multiplication at 0.5 mg/l + 0.5 mg/l Kn in Stevia rebaudiana, Bertoni. Supplemented Kn to the medium enhanced the elongation of shoots.

Figure 2: In vitro regeneration of S. rebaudiana, ‘Bertoni’ a) Initiation and/or establishment of culture in MS supplemented with 0.5 mg/ l BAP b, c) multiplication and elongation of culture in MS supplemented with 1 mg/l BAP + 0.5 mg/l Kn d) rooting of plantlet on ½ strength MS medium supplemented with 2.0 mg/l IBA and e) Hardening of plantlet in pot containing sand andpeat mixture (1: 1).
Root Induction and acclimatization of plant After two cycles of sub-culturing, elongated shoots of 2-3 cm in length were excised and cultured on MS medium having different concentration of IBA. The experiments were conducted twice, with 3 replications (with 3shoots per tubes). Initiation of rooting took place after 1-2 week of inoculation. Single and multiple roots were formed from the base and the nodal portions within 2-3 weeks. The highest number of root (MNR; 9.7±0.45) and root length (MRL; 9.2±0.17) was observed on ½ strength MS medium supplemented with 2.0 mg/l IBA whilelowest number of root (6.4±0.45) and root length (7.2±0.60) occurred in ½ strength MS medium supplemented with 0.5 mg/l IBA (Figure 1c; Figure 2d ). Similar results were observed by Debnath [14]. Abd-Alhady et al. [15] also stated that IBA gave better response for rootingof Periploca angustifolia than NAA.
Plantlets with well-developed root systems were transferred to pots containing sand and peat mixture (1:1) and covered with translucent plastic bags to ensure high humidity around the plants. The use ofthis procedure during the acclimatization phase ensured that most of the plantlets transplanted to ex vitro conditions continued to grow vigorously. After two months when the plastic bags were completelyremoved, 70% of the plantlets survived in the greenhouse and showed normal growth of the plant.
The result of the experiment and other earlier research reports clearly support the possibility of propagating S. rebaudiana, Bertoni, by adopting in vitro techniques. Thus, In vitro propagation can become an important alternative to conventional propagation andbreeding procedures for wide range of plant species.
In vitro salinity and drought stress studies
Fundamentally, plants require energy (light), water, carbon and mineral nutrients for growth. Abiotic stress is defined as environmental conditions that reduce growth and yield below optimum levels. Plant responses to abiotic stresses are dynamic and complex [16]; they are both elastic (reversible) and plastic (irreversible).
Effect of NaCl and mannitol on plant growth Plant growth inhibition is a phenomenon often occurring in glycophyte or saltsensitive plant species under salinity and drought conditions. In the present study, salt and drought stress induced significant differenceson plant growth during the experimental period. The significant decrease in the shoot number and shoot length was observed with the increasing concentration of NaCl (25, 50, 75 and 100mM) and significant increase in shoot number and shoot length with increasing number of days (10, 20 and 30) (Supplementary file 1A; Figure 3a,b;Figure 4). The highest number of shoot (~25) and shoot length (10.7cm) was observed at 25mM after 30 days and lowest number of shoot (~7) and shoot length (6.5cm) at 100mM after 10 days of treatment as compared with control. The plants grown under control(no salinity application) exhibited the maximum values for shootnumber (~28) and shoot length (12.6cm) as compared to remainingtreatments after 30 days of culture, which indicated that salinity isresponsible for reduction in shoot number and shoot length. Shahidet al. [17] observed the reduction in shoot length with increasingconcentration of NaCl. Ashraf et al. [18] also reported that salt stressaffects the plant growth and development by influencing fresh anddry weight of roots, shoots along with shoot length. This might beexplained as the inadequate photosynthesis caused by stomatalclosure and the reduction of carbon assimilation rate under salt stress[6].
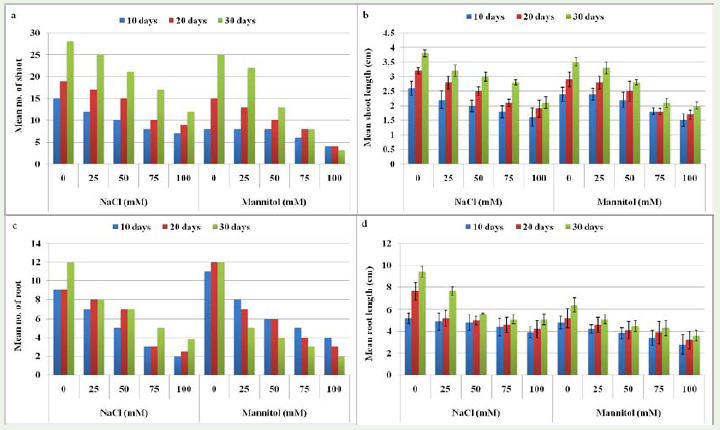
Figure 3: In vitro effect of different concentrations (0, 25, 50, 75 and 100mM) of NaCl and mannitol stress on Stevia rebaudiana ‘Bertoni’ a) Mean number of shoot after 10, 20 and 30 days of culture b) Mean shoot length after 10, 20 and 30 days of culture c) Mean number of root after 10, 20 and 30 days of culture d) Mean root length after 10, 20 and 30 days of culture.

Figure 4: Effect of salinity stress on plant growth and development atdifferent concentrations 25 (a), 50 (b), 75 (c) and 100mM (d) of NaCl.
Drought is a multidimensional stress affecting plants at variouslevels of their organization [19]. The response to drought at the wholeplant and crop levels is complex because it reflects the integration of stress effects and responses at all underlying levels of organizationover time and space. In the present investigation, drought stress exhibited significant differences on shoot number and shoot length during the experimental conditions. After 20 stress days, decreases inshoot number and shoot length was observed in plants grown with 25, 50 75 and 100mM mannitol (Figure 3a, b; Figure 5). In case of shoot number, significant (p < 0.05) differences were not observed with the increasing number of days (Supplementary file 1A). Themeans values of shoot number and shoot length were 16, 14.3, 10.3,7.3, 3.7 and 10.5, 10.3, 9.2, 7.4, 7.1 cm for treatments of 0, 25, 50, 75, and 100mM mannitol, respectively.

Figure 5: Effect of drought stress on plant growth and development atdifferent concentrations 25 (a), 50 (b), 75 (c) and 100mM (d) of mannitol.
Salinity and drought stress also exhibited significant differenceson root number. With the increasing concentration of NaCl andmannitol significant decrease in root number were observed after 10,20 and 30 days. Interestingly, at 100mM Mannitol, decrease in rootnumber 4, 3 and 2 were observed with increasing number of days 10, 20and 30 days, respectively (Supplementary file 1A). Moreover, salinitystress significantly increases the root number after 10, 20 and 30 dayswhile drought stress significantly decreases the root number duringthe experimental condition. Similar result was observed by Abdel-Hussein [20] who evaluated MM106 and Omara apple rootstocks forsalt tolerance in vitro. The reduction in rooting percentage might bedue to the inhibitory effects of salt on the metabolic activities whichassociated with cell division, differentiation and elongation [21].
By following the root length of the plants under salinity anddrought stress during the treatment periods, Fig 3 it appears there isa general trend for the decrease in the root length of the plant usingthe concentration 25, 50, 75 and 100mM while significant increasein the root length after 10, 20 and 30 days. The means values of rootlength were 7.4, 5.9, 5.1, 4.7 and 4.4 cm and 5.5, 4.6, 4.1, 3.9 and 3.2cm for treatments of 0, 25, 50, 75, and 100mM NaCl and mannitol,respectively. Ashraf et al. [18] report that plants grown under control(no salinity application) exhibited the maximum values (10.11cm) for root length as compared to remaining treatments, whichindicated that salinity is responsible for reduction in root length.Thiam et al. [22] reported that root length was adversely affected witha significant reduction of NaCl concentration (50 and 100 mM) and reached 6.95cm and 6.33 cm, respectively. Their findings suggest thathigh NaCl concentration significantly affect the root growth.
Moreover, salinity and drought stress resulting significantdecrease in the leaf FW, leaf DW, stem DW, shoot DW and root DWafter 25, 50, 75 and 100mM treatment. The significant reduction ofleaf FW 36% and 54%, leaf DW 79% and 55%, stem DW 66% and60%, shoot DW 38% and 55%, root DW 72% and 51% were observedat 100mM NaCl and mannitol treatment, respectively. Furthermore,the significant percentage difference in between the salinity anddrought stress conditions were 25%, 21% and 6% of leaf DW, rootDW and stem DW whereas no significant difference were observed inshoot DW and leaf FW (Supplementary file 1B; Table 1, Table 2).
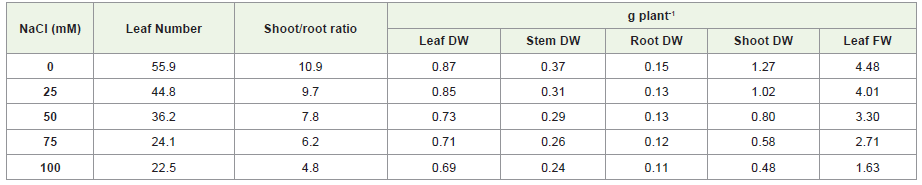
Table 1: Effect of different concentration of NaCl on growth parameters of S. rebaudiana ‘Bertoni’.
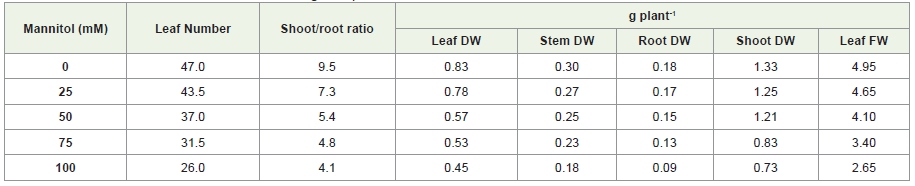
Table 2: Effect of different concentration of mannitol on growth parameters of S. rebaudiana ‘Bertoni’.
Zeng et al. [23] reported that plants treated with 90 and 120 mMNaCl showed significantly lower plant height, leaf number, branchlength, stem dry weight, leaf fresh/dry weight, and shoot dry weightthan control. They also found that high salt treatment (120 mM NaCl)drastically decreased the total dry weight of S. rebaudiana by 40% incomparison to control. In addition, shoot/ root ratios of salt-treatedplants were inhibited markedly by 30-50% compared with that of thecontrol. The results obtained in the present experiment verified thisconclusion that as the salinity increases, the biomass content of S.rebaudiana significantly reduces than control. Srivastav and Srivastav[24] also presented the similar findings that severe drought stressconditions reduce the plant biomass of S. rebaudiana. These resultswere similar to the findings of Pirzad et al. [25] and Mohammadianet al. [26].
Leaf is the main plant organ responsible for light absorptionand transformation and plays an important role in providing energyand nutrients for the growth and development of plants. The growthinhibition caused by salt and drought stress mainly originated fromthe damage to leaf function. Results presented in Table 1 and Table 2 showed that higher levels of salinity and drought decrease leafnumber throughout the experiment. In the present study, the numberof fully expanded leaves decreased dramatically with increasingsalt level, which could be seen as the combined effect of increasingthe senescence rate of older leaves and decreasing the productionof new leaves. Zeng et al. [23] reported leaf chlorosis, wilting, andnecrosis were in plants treated with 90 and 120 mM NaCl for 28 days,respectively. It was found that the general trend of the treatmentreflects a gradual decrease in the number of plant leaves with the increase of NaCl and mannitol concentration, compared with theplants of the control experiment.
Leaf number significantly reduced as increasing concentrationof NaCl and mannitol. It was found that the general trend of thetreatment reflects a gradual decrease in the number of plant leaveswith the increase of salt concentration, compared with the plantsof the control experiment. The maximum number of leaves 44.8and 43.5 were found at 25mM and minimum number of leaves 22.5and 26 at 100mM NaCl and manitol stress treatments, respectively.According to Silva et al. [27] leaves were more sensitive than plantheight after 7 days of treatment. After 36 days, significant reductionin leaves number at all NaCl levels was observed. Shao et al. [28]reported that reduction in number of leaves under water stress ismay be due to decline in cell enlargement and more leaf senescenceresulting from reduced turgor pressure. On the other hand, significantreduction in shoot/root ratio was also observed as NaCl and manitolconcentration increase (Table 1). Shoot growth was probably moresusceptible to salt than root, which generally led to a lower shoot/rootratio in salt treatments. Zeng et al. [23] found significant reduction inshoot/root ratios in all NaCl treatments than in control. Moreover,Nejad [29] studied on effect of drought stress on shoot/root ratio withfour levels of water stress as the first factor and plant growth periodsas the second factor with four replications. Their findings suggestthat increasing drought stress with each period of growth, shoot androot weight decreases per plant. Previous studies suggested that thissensitivity could be due to an imbalance among cations as a result ofthe complex interaction in the xylem transport system.
Effect of salinity and drought stress on protein content Figure 6b and Figure 7b results indicate a reverse effect of sodium chlorideand mannitol using various concentrations on total protein of S.rebaudiana plant after 10, 20 and 30 days. It appears from the datathat there was a general decrease in protein content that correspondedwith the increase in NaCl and mannitol concentrations as comparedwith the control plant. Statistical analysis did show a significantdifference during the three measurement periods (Supplementary file 2A, B).
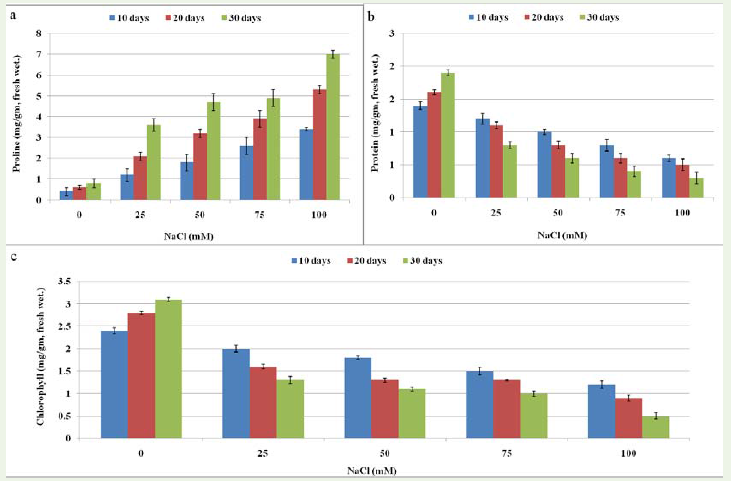
Figure 6: Effect of different concentrations (0, 25, 50, 75 and 100mM) of NaCl on biochemical content of Stevia rebaudiana ‘Bertoni’ a) Effect of salinity stress on proline content of plant b) Effect of salinity stress on protein content and c) Effect of salinity stress on chlorophyll content.
Chen et al. [30] found that exposing Vigna unguiculata (L.) plants,at the age of 14 days, to salt treatment using 75mM of NaCl, reducedsoluble protein content in the plant. The results of the previous studywas confirmed by Cheruth et al. [31], with their study on Cathranthusroseus (L.) and Khosravinejad et al. [32] with their study on barley Hordeum vulgare L. On the contrary, Sibole et al. [33] reported thatthe treatment of clover plant (Medicago citrna L.) for 30 days withconcentrations of 0, 1, 50, 100, 200 mM of NaCl increased solubleprotein content in the seedlings, compared with control plants.Another study by Kapoor and Srivastava [34] on Vigna mungo (L.)supports the previous results.
In contrast to effect of drought stress on protein concentration,there were no significant differences observed with the increasingconcentration of mannitol as compared to control plants. However,the amount of protein (mg/gm) increases after 10, 20 and 30 daysof culture in control plant. Several researches indicated that solubleprotein decreases with increasing drought stress time [35,36]. Thewater stress injury causes damage to protein synthesizing mechanism.The possible reason for decreased protein content under water stressmay be due to increased activity of protease and also it may be dueto proteolysis or decreased synthesis or both. Leaf proteins undergoaccelerated hydrolysis as water stress develops.
Effect of salinity and drought stress on proline content We analyzedthe proline content of plant grown at 0, 25, 50, 75 and 100mM NaCland mannitol. Results showed that amount of proline increased withthe increasing concentration of NaCl and mannitol (Figure 6a; Figure 7a). Statistically significant differences in proline accumulation weredetermined between the control and treatment periods of NaCl whilein drought stress condition no significant differences were observed(Supplementary file 2A, B). Proline accumulation in salt stressedplants is a primary defense response to maintain the osmotic pressurein a cell, which is reported in salt tolerant and salt sensitive cultivarsof many crops [37-39]. Zhang et al. [23] reported that proline contentenhanced significantly with increasing concentration of NaCl andwas 17-, 31-, and 42-fold higher in 60, 90, and 120 mM NaCl than inthe control, respectively. Several studies have indicated that prolinecontent increases during drought stress [40,41].
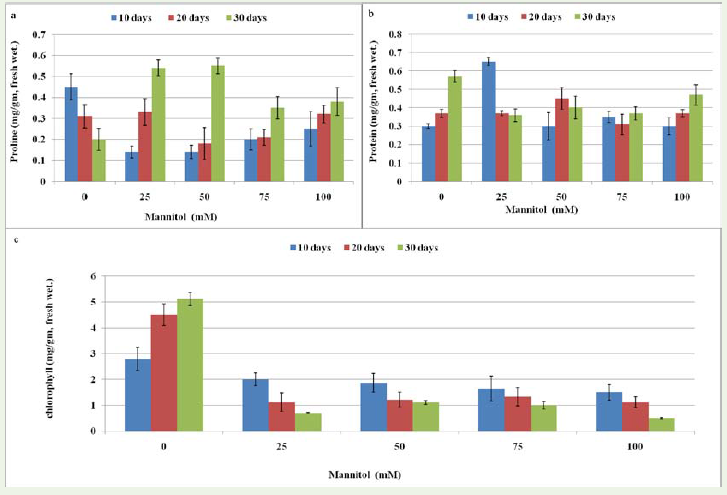
Figure 7: Effect of different concentrations (0, 25, 50, 75 and 100mM) of mannitol on biochemical content of Stevia rebaudiana ‘Bertoni’ a) Effect of drought stress on proline content of plant b) Effect of drought stress on protein content and c) Effect of drought stress on chlorophyll content.
Effect of salinity and drought stress on chlorophyll content Decrease in chlorophyll content was determined as a result of increasing NaCl and mannitol concentration. The highest content of chlorophyll was found at 25mM after 10 days of culture while lowest content wasfound at 100mM after 30 days in both salinity and drought treatment conditions. The significant decreasing percentage of chlorophyll was 50 % at 100mM NaCl treatment after 10 days of culture that further reduced (16 %) after 30 days of stress treatment. Moreover, significantreduction (54 %) was observed at 25mM after 100 days of mannitol that decrease (10 %) further after 30 days of culture (Figure 6C; Figure 7C). Statistical analysis shows that significant reduction in chlorophyll content was found at each concentration level (25, 50, 75 and 100mM)but data obtained after 10, 20 and 30 days of culture was found no significant in both salinity and drought stress plants (Supplementary file 2A, B). The decrease in chlorophyll content may be due to an increase of chlorophyll degradation or to a decrease of chlorophyllbiosynthesis. Change in chlorophyll contents due to salinity is the most evident biochemical response [42]. Amira et al. [43] also found the similar results on propagation of plant Cathranthus roseus under salinity and drought conditions where they reported that reduction in chlorophyll associated with reduced photosynthetic activity andtranspiration rate in plants at all salinity levels is considered a defense mechanism against damaging reactive oxygen species by diminishing light absorbing capacity that reduces the flow of electrons throughthe photosysteme. Moreover, disruption of the thylakoid and stromal membranes may result in leaf chlorosis and necrosis, Qureshi et al. [44] which indicated that salt concentration above 90 mM NaCl could impair the chloroplast structure of S. rebaudiana. In contrast to drought stress condition, decrease in the chlorophyll was observed which is reported first time in S. rebaudiana. No such study was previously reported in S. rebaudiana.
Summarizing, salinity and drought stress significantly affect the growth and development as well as biochemical content of S. rebaudiana. As the concentration of NaCl and mannitol increased physiomorphologic and biochemical components i.e. proline, protein and chlorophyll contents negatively affects the growth and development of plants but 50 % of plant was survived at the concentration (50mM) of NaCl and mannitol. These results indicatethe mutant development at such concentration. Further research is in progress to decipher the expression profile of four major genes involved in steviol glycoside pathway under salinity and drought conditions and compare with HPLC results for stevioside and rebaudioside i.e. responsible for sweetening property, content withcontrol plant.
Acknowledgment
The authors would like to thank to the Director of Xcelris Genomics for providing facilities and assistance.
References
- Bray EA, Bailey-Serres J, Weretilnyk E (2000) Responses to abiotic stresses. In: Gruissem W, Buchannan B, Jones R (eds) Biochemistry and Molecular Biology of Plants. American Soc Plant Physiol, Baltimore, MD: 1158-1249.
- Lemus-Mondaca R, Vega-Galvez A, Zura-Bravo L, Ah-Hen K (2012) Stevia rebaudiana Bertoni, source of a high-potency natural sweetener: a comprehensive review on the biochemical, nutritional and functional aspects. Food Chem 132: 1121-1132.
- Liu J, Li SFY (1995) Separation and determination of Stevia sweeteners by capillary electrophoresis and high performance liquid chromatography. J Liquid Chromatography 18: 1703-1719.
- Pol J, Ostra EV, Karasek P, Roth M, Benesova K, et al. (2007) Comparison of two different solvents employed for pressurised fluid extraction of stevioside from Stevia rebaudiana: methanol versus water. Anal Bioanal Chem 388: 1847-1857.
- Felippe GM, Lucas NMC (1971) Estudo da viabilidade dos fructos de Stevia rebaudiana Bert. Hoehnea 1: 95-105.
- Ben Ahmed C, Ben Rouina B, Sensoy S, Boukhriss M, Abdullah FB (2009) Saline water irrigation effects on antioxidant defense system and proline accumulation in leaves and roots of field-grown olive. J Agric Food Chem 57: 11484-11490.
- Sheng RL, Yuan HY, Huang SZ (2011) Influences of Na2CO3 stress on physiological metabolisms of different alkali tolerant varieties of Stevia rebaudiana. Agric Sci Technol 12: 1837-1841.
- Debnath M, Pandey M, Chikara, SK (2011) Physiological and molecular characterization of in vitro cultures of an endemic medicinal herb, Chlorophytum borivilianum, under abiotic stress. African J Biotechnol 10: 7356-7366.
- Bates LS, Waldren RP, Teare ID (1973) Rapid determination of free proline for water-stress studies. Plant Soil 39: 205-207.
- Hurkman WJ, Tanaka CK (1986) Solubilization of plant membrane proteins for analysis by two-dimensional electrophoresis. Plant Physiol 81: 802-806.
- Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951) Protein measurement with the folin phenol reagent. J Biol Chem 193: 265-275.
- Porra RJ, Thompson WA, Kriedemann PE (1989) Determination of accurate coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: verification of the concentrarion of chlorophyll standards by atomic absorption spectroscopy. Biochem Biophys Acta 975: 384-394.
- Abd-Alhady MRA (2011) Micropropagation of Stevia rebaudiana Bertoni. A New Sweetening Crop in Egypt. Global J Biotechnol Biochem 6:178-182.
- Debnath M (2008) Clonal propagation and antimicrobial activity of an endemic medicinal plant Stevia rebaudiana. J Med Plant Res 2: 045-051.
- Abd-Alhady MRA, Abd Alla MM, Hegazi GA, Gabr MF (2010) Rapid propagation of Periploca angustifolia Labill. By tissue culture. International J Plant Developmental Biol 4: 15-18.
- Cramer GR (2010) Abiotic stress & plant responses from the whole vine to the genes. Aust J Grape Wine Res 16: 86-93.
- Shahid MA, Pervez MA, Balal RM, Ahmad R, Ayyub CM, et al. (2011) Salt stress effects on some morphological and physiological characteristics of okra (Abelmoschus esculentus L.). Soil Env 30: 66.
- Ashraf M, Arfan M, Ahmad A (2003) Salt tolerance study in okra: ion relations and gas exchange characteristics. J Plant Nutr 26: 63-79.
- Wentworth M, Murchie MH, Gray JE, Villegas D, Pastenes C, et al. (2006) Differential adaptation of two varieties of common bean to abiotic stress. II. Acclimation of photosynthesis. J Exp Bot 57: 699-709.
- Abdel-Hussein MA-A (2006) Evaluation of MM106 and. Omara apple rootstocks for salt tolerance in vitro. J Karbala Univ 4: 184-191.
- Binh DQ, Heszky LE, Gyulai G, Csillag A (1992) Plant regeneration of NaCl-pretreated cells from long-term suspension culture of rice ( Oryza sativa L. ) in high saline conditions. Plant Cell Tiss Org Cult 29: 75-82.
- Thiam M, Champion A, Diouf D, Mame Oureye SY (2013) NaCl Effects on In Vitro Germination and Growth of Some Senegalese Cowpea (Vigna unguiculata (L.) Walp.) Cultivars. ISRN Biotechnol: 11.
- Zeng J, Chen A, Li D, Yi B, Wu W (2013) Effects of Salt Stress on the Growth, Physiological Responses, and Glycoside Contents of Stevia rebaudiana Bertoni. J Agric Food Chem 61: 5720-5726.
- Srivastava S, Srivastava M (2014) Influence of water stress on morpho-physiological and biochemical aspects of medicinal plant Stevia Rebaudiana. Life Sciences Leaflets 49: 35-43.
- Pirzad A, Khoshbakht M, Siadat AA, Ghodrat AF, Bakhshande AM (2012) Growth analysis of Pimpinella anisum under different irrigation regimes and amounts of super absorbent polymer. Inter Res J App Bas Sci 3: 112-122.
- Mohammadian R, Moghaddam M, Rahimian H, Sadeghian SY (2005) Effect of early season drought stress on growth characteristics of sugar beet genotypes. Turk J Agric For 29: 357-368.
- da Silva EC, Nogueira RJMC, de Araujo FP, de Melo NF, de Azevedo Neto AD (2008) Physiological responses to salt stress in young umbu plants. Environ Exp Bot 63: 147-157.
- Shao HB, Chu LY, Jaleel CA Zhao CX (2008) Water-deficit stress-induced anatomical changes in higher plants. C R Biologies 331: 215-225.
- Nejad TS (2011) Effect of Drought Stress on Shoot / Root Ratio. World Academy of Science, Engineering and Technology 5: 539-541.
- Chen Z, Cuin TA, Zhou M, Twomey A, Naidu BP, Shabala S (2007) Compatible solute accumulation and stress-mitigating effects in barley genotypes contrasting in their salt tolerance. J Exp Bot 58: 4245-4255.
- Cheruth AJ, Ragupathi G, Ashot K, Paramasivam M, Beemarao S, et al. (2008) Interactive effects of triadimefon and salt stress on antioxidative status and ajmalicine accumulation in Catharanthus roseus. Acta Physiol Plant 30: 287-292.
- Khosravinejad F, Heydari R, Farboodnia T (2009) Effect of salinity on organic solutes contents in barley. Pak J Biol Sci 12: 158-162.
- Sibole JV, Cabot C, Poschenreder C, Barcelo J (2003) Efficient leaf ion partitioning, an overriding condition for abscisic acid-controlled stomatal and leaf growth responses to NaCl salinization in two legumes. J Exp Bot 54: 2111-2119.
- Kapoor K, Srivastava A (2010) Assessment of salinity tolerance of Vinga mungo var. Pu-19 using ex vitro and in vitro methods. Asian J Biotechnol 2: 73-85.
- Guangxi R, Xiangyang L, Yan S (2011) Effects of Drought Stress on Physiological Indices and Dry Leaf Yield of Stevia rebaudina Bertoni. J Agricult: 11.
- Surendar KK, Devi DD, Ravi I, Jeyakumar P, Velayudham K (2013) Water Stress Affects Plant Relative Water Content, Soluble Protein, Total Chlorophyll Content and Yield of Ratoon Banana. Inter J Hort 3: 96-103.
- Kumar SG, Reddy AM, Sudhakar C (2003) NaCl effects on proline metabolism in two high yielding genotypes of mulberry (Morus alba L.) with contrasting salt tolerance. Plant Sci 165: 1245-1251.
- Misra N, Gupta AK (2005) Effect of salt stress on proline metabolism in two high yielding genotypes of green gram. Plant Sci 169: 331-339.
- Veeranagamallaiah G, Chandraobulreddy P, Jyothsnakumari G, Sudhakar C (2007) Glutamine synthetase expression and pyrroline-5-carboxylate reductase activity influence proline accumulation in two cultivars of foxtail millet (Setaria italica L.) with differential salt sensitivity. Environ Exp Bot 60: 239-244.
- Ghorbanli M, Gafarabad M, Amirkian T, Allahverdi MB (2013) Investigation of proline, total protein, chlorophyll, ascorbate and dehydro ascorbate changes under drought stress in Akria and Mobil tomato cultivars. Iranian J Plant Physiol 3: 651-658.
- Mafakheri A, Siosemardeh A, Bahramnejad B, Struik PC, Sohrabi Y (2010) Effect of drought stress on yield, proline and chlorophyll contents in three chickpea cultivars. Australian J Crop Sci 4: 580-585.
- Jamil M, Bashir S, Anwar S, Bibi S, Bangash A, et al. (2012) Effect of salinity on physiological and biochemical characteristics of different varieties of rice. Pak J Bot 44: 7-13.
- Amira MH, Mohamed EHO, Sayed MH, Soad SE (2007) Effect of salinity and drought on growth criteria and biochemical analysis of Cathranthus roseus shoot. Int J Bot 3: 202-207.
- Qureshi IM, Israr M, Abdin MZ, Iqbal M (2005) Responses of Artemisia annua L. to lead and salt-induced oxidative stress. Environ Exp Bot 53: 185-193.