Modern Pollen Assemblages of Surface Samples from Tropical Deciduous Forest of Assam, Northeast India: A Window to Palaeoclimatic Interpretation
Swati Tripathi * and S.K. Bera
Corresponding author: Swati Tripathi, Quaternary Palynology Division, Birbal Sahni Institute of Palaeobotany, 53University Road, Lucknow-226007, Uttar Pradesh, India; E-mail: swatidixit26@gmail.com
Quaternary Palynology Division, Birbal Sahni Institute of Palaeobotany, 53 University Road, Lucknow-226007, Uttar Pradesh,India
Citation: Swati T, Bera SK. Modern Pollen Assemblages of Surface Samples from Tropical Deciduous Forest of Assam, Northeast India: A Window to Palaeoclimatic Interpretation. J Plant Sci Res. 2014;1(2): 108.
Copyright © 2014 Swati Tripathi et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Plant Science & Research | ISSN: 2349-2805 | Volume: 1, Issue: 2
Submission: 29/05/2013; Accepted: 03/07/2014; Published: 05/07/2014
Reviewed & Approved by: Dr. Firoze Quamar, Scientist, Birbal Sahni Institute of Palaeobotany, India
Abstract
Palynological assessment of 20 surface samples from the Panchratna reserve forest of Goalpara District, western Assam has been carried out to explore the relationship between modern pollen assemblages and contemporary vegetation patterns. Seven spider web samples have also been examined in order to conceive air pollen-spore and its relation to soil/moss pollen data. The surface samples have depicted the occurrence of mixed pollen assemblage comprising Angiosperms, Gymnosperms and Pteridophytes among which non-arboreals (terrestrial herbs and marshy/aquatic taxa) dominate over arboreals (trees and shrubs). The comparison between pollen spectra and vegetation reflects site to site variation in pollen assemblage in relation to heterogeneity of the forest. The occurrence of highland plants is suggestive of long distance transportation of pollen from higher elevation. The affluence of both monolete and trilete fern spores indicate humid climatic condition. Pollen frequencies of surface samples somehow portray the extant floral distribution in and around the reserve forest which is well evidenced from our palynological data where relative frequency of the assemblage includes local arboreals at the average values of 33.91%, 22.88% and 11.33% at centre, margin and open-land area of the forest respectively. Thus, the surface samples from near proximity of the woods dominate in the arboreal pollen frequencies which are quite relevant to extant floral composition. The representation of fungal elements belongsto Deuteromycetes and Ascomycetes (grass pathogen) as evidenced by degraded pollen and spores which are suggestive of biological degradation in sediments. Therefore, there is a need to precisely observe the behavioural pattern of modern pollen deposition which could be helpful in deciphering past climate and vegetation in and around the pristine floodplain ecosystem of Northeast India which is at the verge of deterioration.
Keywords: Pollen rain; Tropical deciduous forest; Goalpara district; Northeast India
Introduction
The characterisation of modern ecosystems by their pollen rain is an essential prerequisite for interpreting Late Quaternary fossil pollen data [1]. The paucity of modern pollen-rain studies from the tropics of India is therefore a significant hindrance to resolving queries concerning the Late Quaternary vegetation and climate succession of the country [2-4]. Moreover, due to the considerable heterogeneity of forest ecosystems across the Indian subcontinent [5], one cannot assume that the pollen-rain signatures of these isolated study sites are necessarily representative of forests in other parts of the country, as highlighted by the dissimilarity between forest pollen-rain signatures of tropical deciduous Sal forest of northeast India compared with those from Sal forest of central India [6,7]. Qualitative studies of pollen-vegetation relationship in several vegetation types in Indiansubcontinent have been carried out [8-10]. However, only a few studies concerned the quantitative relationship between pollen and vegetation for example [11,12].
Pollen and spores are the basis of an important aspect of palaeoclimatic reconstruction, generally referred to as palynology, the study of spores and pollen. Where pollen and spores have accumulated over time, a record of the past vegetation of an area may be preserved. In many cases changes in the vegetation of an area may be due to changes in climate; hence, interpreting past vegetation through palynology may lead to strong inferences about the former climate of an area. Ofcourse, not all changes in the accumulation of pollen and spores are necessarily due to change in climate; fire, insect infestation, plant successional changes, interference by man as well as changes in factors leading to the accumulation and preservation of the fossil material itself often makes interpretation of the pollen and spore record complex. The pollen analyst must therefore carefully evaluate the record to isolate whatever climate signals it contains. A detailed palaeoclimate record, spanning at least the late Quaternary period, is therefore fundamental to comprehension of modern climate, and the causes of climatic variation and change [13]. Furthermore, unless the natural variability of climate is understood, it will be extremely difficult to recognize any man-made effects on climate that may become apparent in the future [14].
Northeast India forms a significant portion of both the Himalaya and Indo-Burma biodiversity hotspots [15]. We present here modern pollen assemblage data of 20 sites from Panchratna reserve forest of Goalpara district, Assam. This study provides detailed information on vegetation composition and modern pollen assemblage from the centre, margin and open-land area of the forest. Our objective is to understand the relationship of modern pollen assemblage to the source of vegetation and regional climate in the study area based on pollen ratio calculations, in order to provide a basis for the reconstruction of palaeovegetation and palaeoclimate. In addition past vegetational shifts can provide a clue to rapid climatic changes incyclic/random pattern and also help in tracing the sequence and time of beginning of deterioration of reserve forests in Assam.
However, the forests distributed in Assam are under tremendous pressure of exploitation. Studies have indicated that with in a period of two years between 1995 to 1997, about 1,875 sq km of forest land were converted to crop land for shifting cultivation, whereas, the natural regeneration was recorded over only 1,700 sq km of land resulting in a net loss of 175 sq km of forest over a two-years period [16]. The state of Assam bordered on north by Bhutan and Arunachal Pradesh, on east Arunachal Pradesh, Nagaland and Manipur, on south Mizoram, and on east by Tripura, Bangladesh, Meghalaya and West Bengal. Thus, represents a unique physiographic makeup characterized by hills, plateau and flood plains. The hills are extensions of Himalayan mountain range and geological of recent (Tertiary) formation. The plains namely Brahmaputra and Barak are formed as a result of deposition of alluvial sediments on the Tertiary and Mesozoic bed rocks and the depressions are developed due to tectonic disturbances [17].
Hitherto, not much study have been performed concerning modern pollen/vegetation relationship so far from reserve forests of Assam except few publications from Dabaka, Chayagaon, Dibrugarh and Kaki forest division [18,19,7,20]. Panchratna reserve forest is located on the southern bank of the river Brahmaputra situated at a distance of about 9 km west of Goalpara District (Figure 1). The total area of this reserve forest is 976.120 hectares. The reserve forest comprises of predominantly hilly terrain associated with a number of swampy areas. Goalpara District is situated on the south bank of river Brahmaputra. The District covers an area of 1,831 sq km and is bounded by west and east Garo Hills, Meghalaya on the south, Kamrup District on the east, Dhubri District on the west and river Brahmaputra all along the north. The geographical location of the District is between latitude 25°92’ to 26°07’ N and longitude 90°01s’ to 91°15’ E. The topography of Goalpara District is generally characterized by cultivable plains, except for few low-forested Hill regions. The main tributaries of the Brahmaputra River flowing through the District are Dudhnai, Krishnai, Balbala, Jinjiram, Jinari and Deosila. All the rivers are perennial in nature. Natural lakes such as Urpad Beel, Hasila Beel, Kumuri Beel and Dhamar Risan Beel exist in the District and several other artificial ponds are also seen at places. There are about 165 small and large wetlands covering an area of 3,832.50 hectares along with number of reserve forests in the District. The District is primarily agrarian as 90% of the population depends for their livelihood on agriculture. The District is industrially backward and there are no existing industries. It was a land locked District with poor transport and communication facilities till the opening of rail cum Road Bridge, the Namonarayan setu, over the river Brahmaputra.
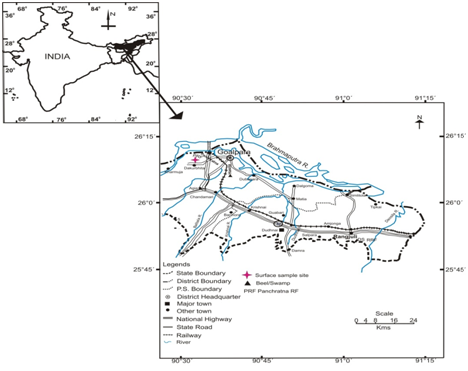
Figure 1: Location map showing study site.
Geology and Climate
Older alluvium in the Goalpara District is covered by recent alluvium throughout its stretches. The drainage pattern in the valley is of antecedent type. The often changing curvy courses of the Brahmaputra River and its tributaries are not only due to lateral erosion because of the low gradient of the rivers but also due to periodic, local and sudden changes in the basement levels due to the earthquakes. The soil in the study area is characterized by recent deposition of alluvium, moderately deep to very deep with grey to molted grey colour (Table 1). It is mostly composed of sandy to silty loams and slightly acidic in nature [21]. On the riverbanks, it is less acidic and sometimes neutral or slightly alkaline. The soil lack in profile development and is deficient in phosphorus, nitrogen and humus. The climate in the District is moderately cold during the winter and in summers, it is hot. Rainfall makes its first appearance in April with occasional and irregular light showers and at times heavy downpour followed by cyclonic storms. The irregular rainfall continues up to the end of May. This rain occurs due to the influence of North-Eastern wind. Monsoon rain normally begins from the early part of June and heavy rains occur in the District till October. About 80% rainfall is from South West monsoon. The maximum temperature is 36°C during July and August, whereas, minimum temperature dips down to 7°C in the month of January at places (http:/www.britannica.com/EBchecked/topic/39101/Assam).
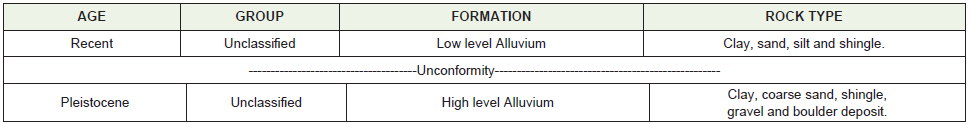
Table 1: The Quaternary geological formations covering the state of Assam are classified into the following order of sequence (Karunakaran, 1974).
The study area, in general, experiences a warm and humid climate, which is largely influenced by southwest monsoon. The mean minimum and maximum winter temperatures are 16.3°C and 21°C, respectively, however, the lowest temperature of 1°C is recorded during the month of January. The mean minimum and maximum summer temperatures are 31.6°C and 34°C, respectively. During the extreme hot summer month of June the temperature occasionally rises up to 44°C. The rainfall by and large falls from mid June to September, however, major fraction of it takes place in July and August. The average annual precipitation recorded for the area is 1200mm. Approximately 92% of the total precipitation occurs during the rainy season.
Vegetation of the study site
The vegetation is generally characterized by tropical mixed deciduous forest comprising prominent trees like Shorea robusta, Terminalia bellirica, Dillenia pentagyna, Artocarpus chaplasha, Lannea coromandelica, Careya arborea, Semecarpus anacardium, Sterculia villosa, Sapindus rarak, Combretum latifolium, Syzygiumcumini, Schima wallichii, Ilex sulcata, Emblica officinalis, Sterculia villosa, Salmalia malabaricum Lagerstroemia speciosa and L. parviflora (Figure 2a). However, Arecaceae and wild bananas (Musa spp.) form a regular stand at the hilly slope (Figure 2b). Shrubs such as Melastoma malabathricum, Bauhinia purpurea, Justicia adhatoda, Holarrhena pubescens, Leea crispa and Strobilanthes scaber are represented as dominant shrubby taxa. The bamboo stakes are composed of Dendrocalamus hamiltonii, Bambusa pallida, B. polymorpha, B. vulgaris, Dinochloa compactiflora and Melocanna bambusoides at places adjacent to the vast swamps. The grasses along with a large number of terrestrial herbs comprise the ground flora. The major taxa are Xanthium strumarium, Artemisia parviflora, Verbenaceae, Caryophyllaceae, Amaranthaceae, Chenopodiaceae, Tubuliflorae, Liguliflorae, Brassicaceae, Lamiaceae, Euphorbia hirta and Cannabis sativus. The fern allies like Lycopodium clavatum, Angiopteris evecta, Dryopteris filix-mas, Cyathea gigantea, Lygodium japonicum, Pteris vittata, P. stenophyllum, P. biaurita, P. cretica and Asplenium cheilosorum are occurred frequently usually along road cutting.

Figure 2: a) View showing tropical mixed deciduous forest of Panchratna, Goalpara, Assam, b) View showing regular stand of Arecaceae and wild bananas (Musa spp.) at the hilly slope along with cultivable plain.
Materials and Methods
Field work
We collected vegetation data and subsurface soil samples from 20 sites from vegetation zones with different degrees of degradation. About 46 important polliniferous materials were procured from in and around the study area for understanding the pollen morphological details [22]. The location of each pollen site was determined by GPS. Samples were collected in a transect from within forest across openland area at a distance of 50 m interval. Of the 20 samples, 8 were procured from the vicinity of the swamp at the centre of the reserve forest, 6 each from forest edge and open land area to study modern pollen/vegetation relationship. For the present investigation the spider webs were collected during winter season (October-December, 2008). The sample collection and chemical processing of spider webs was adopted from the earlier published research paper [23]. The detail location and vegetation information from the regional surface pollen sample sites are mentioned in Table 2.
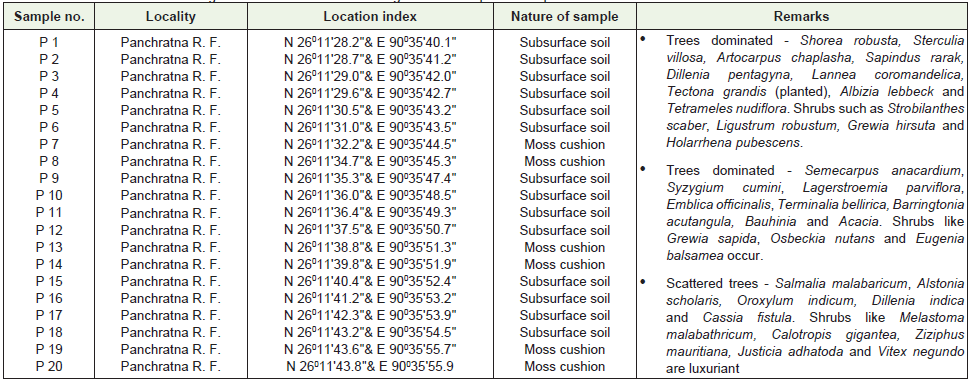
Table 2: The detail location and vegetation information from the regional surface pollen sample sites.
Laboratory work
The samples were processed employing standard acetolysis method [24] as the collected samples were treated with 10% aqueous KOH solution to deflocculate the pollen/spore from the soil samples followed by 40% HF treatment to dissolve silica content. Thereafter, the conventional procedure of acetolysis was followed using acetolysis mixture (9:1 acetic anhydrite and conc. H2SO4). Finally the material was kept in 50% glycerin solution. A few drops of phenol were also added to the glycerin solution to protect the maceral from microbial decomposition [24]. 300 to 500 pollen and spore per sample were counted to make pollen spectra. Plant elements in the study have been categorized into arboreals (trees and shrubs), non-arboreals (terrestrial herbs and marshy/aquatics), ferns and highland taxa. Fungal remains were excluded from pollen sum. For the precise identification of fossil palynomorphs in the soil samples, the reference pollen slides available at BSIP Herbarium as well as the pollen photographs in the published literature [25,26] were consulted. The pollen spectra were made using Microsoft excel programme and modified in Corel Draw-12 software. The frequency percentage of the recovered palynomorphs has been calculated in terms of total pollen and spore count (Figure 3).

Figure 3: a) Pollen spectra from Panchratna reserve forest, Goalpara, Assam.
Modern Pollen Rain
Pollen spectra (Within forest): Sample nos-P 1-6 (SS) and P 7-8 (MC): The palynological study of subsurface soil and moss cushion shows that an average value of non-arboreal taxa (57.36%) is predominant over arboreal taxa (33.91%), ferns (5.97%) and highland taxa (2.76%). The major non-arboreal taxa are represented by grasses including Poaceae<60μm (13.96%) and Cerealia>60μm (6.20%) attaining upto a total of 20.16% along with other terrestrial herbs like Tubuliflorae (3.19%), Lamiaceae, Acanthaceae and Xanthium (2.66% each), Liguliflorae (2.44%), Amaranthaceae and Caryophyllaceae (2.31% each) in moderate values. Plantago (1.10%) and Mimosa (1.08%) are observed in lower values, however, Artemisia is sporadic.The marshy taxa are represented by moderate values of Rumex nepalensis (2.66%) and Colocasia esculenta (2.00%) followed by low values of Onagraceae and Impatiens balsamina (1.74% each) and Cyperaceae (1.61%) with trace values of Polygonum plebeium (1.13%) and P. serrulatum (1.08%). Aquatic taxa are represented by Nymphoides indica (1.74%) and Lemna minor (1.10%) in lower values. Rest of the aquatics like Potamogeton pectinatus, Nymphaea nouchali and Typha latifolia are sporadic. Among ferns, monolete (4.90%) and trilete spores (1.07%) are encountered in high to low value respectively.
Among arboreals, tree taxa like Terminalia bellirica (2.94%), Dillenia pentagyna (2.75%), Lagerstroemia parviflora (2.02%), Shorea robusta (2.00%), Meliaceae (1.95%), Emblica officinalis and Syzygium cumini (1.92% each) are found in moderate values. Whereas, Sapotaceae (1.68%), Adina cordifolia (1.49%), Lannea coromandelica and Semecarpus anacardium (1.42% each) and Salmalia malabaricum(1.04%) are encountered in lower values. Other deciduous tree taxa like Acacia catechu, Albizia lebbeck and Arecaceae are sporadic. Semi evergreen tree taxa like Ilex sulcata (1.90%), Schima wallichii (1.68%), Artocarpus chaplasha (1.42%) and Symplocos racemosa (1.01%) are also encountered in lower values. Among prominent shrubs, Ziziphus mauritiana (2.09%) is found in moderate value along with low valueof Oleaceae (1.71%) whereas, Melastoma malabathricum is sporadic. Highland taxa are represented by Pinus, Betula and Alnus within 2.76%. Fungal remains are also encountered in fairly good values and are represented by Microthyriaceae, Xylaria, Cookeina, Alternaria, Nigrospora, Diplodia and Circinoconis.
Pollen spectra (Forest edge): Sample nos-P 9-12 (SS) and P 13-14 (MC): The palynological study of subsurface soil and moss cushion shows that an average value of non-arboreal taxa (68.13%) is dominant over arboreal taxa (22.88%), ferns (5.64%) and highland taxa (3.35%). The major non-arboreal taxa are represented by grasses including Poaceae<60m (12.34%) and Cerealia>60μm (10.65%) attaining upto a total of 22.99%. Other major terrestrial herbs are recognized by Tubuliflorae (4.72%), Liguliflorae, Amaranthaceae and Caryophyllaceae (3.50% each), Lamiaceae, Acanthaceae and Xanthium (2.23% each), Plantago (2.00%) and Mimosa pudica (1.98%)at moderate values along with low value of Artemisia (1.70%). Marshy taxa are represented by Cyperaceae (3.21%) at moderate value along with low values of Polygonum plebeium, P. serrulatum and Colocasia esculenta (1.65% each), Rumex nepalensis (1.45%), Onagraceae and Impatiens balsamina (1.21% each). Aquatic taxa are represented by Nymphaea nouchali and Potamogeton pectinatus (1.45% each) andNymphoides indica (1.21%) in lower values, however, Typha latifolia and Lemna minor are found sporadic in the assemblage. Among ferns, monolete (4.23%) and trilete spores (1.41%) are observed in high to low value respectively.
Among arboreals, tree taxa include Terminalia bellirica (2.43%) followed by Dillenia pentagyna (2.25%), Lagerstroemia parviflora (2.02%) in moderate values whereas, Sapotaceae (1.68%), Lannea coromandelica and Semecarpus anacardium (1.42% each), Shorea robusta (1.40%), Emblica officinalis and Meliaceae (1.07% each),Syzygium cumini and Adina cordifolia (1.02% each) are found in low values. Rest of the deciduous tree taxa namely Salmalia malabaricum, Acacia catechu, Albizia lebbeck and Arecaceae are sporadic. Semi evergreen tree taxa are represented by Ilex sulcata (1.05%) and Artocarpus chaplasha (1.00%) in lower values whereas, Schimawallichii and Symplocos racemosa are encountered sporadically in the assemblage. Among prominent shrubby taxa, Melastoma malabathricum (1.20%) is predominant over Oleaceae and Ziziphus mauritiana. Highland taxa are represented by Pinus, Betula and Alnus within the value of 3.35%. Fungal remains are encountered inmoderate values as compared to preceding case.
Pollen spectra (Open-land): Sample nos-P 15-18 (SS) and P 19-20 (MC): The palynological study of subsurface soil and moss cushion shows that an average value of non-arboreal taxa (75.91%) is dominant over arboreal taxa (11.33%), ferns (6.05%) and highland taxa (6.71%). The major non-arboreal taxa are represented bygrasses including Cerealia>60μm (15.23%) and Poaceae < 60μm (4.88%) attaining upto a total of 20.11%. Other terrestrial herbs are encountered as Tubuliflorae (6.05%), Plantago (3.08%), Liguliflorae (2.88%), Lamiaceae, Acanthaceae and Xanthium (2.66% each), Amaranthaceae and Caryophyllaceae (2.31% each) at moderately high values followed by Artemisia (1.80%) and Mimosa pudica (1.75%) in lower values. Marshy taxa are represented by Cyperaceae (4.52%), Rumex nepalensis (3.08%), Colocasia esculenta (2.66%) and Polygonum plebeium (2.24%) at moderate values followedby low values of Polygonum serrulatum (1.87%), Onagraceae and Impatiens balsamina (1.76% each) at moderate values. Aquatic taxa are represented by moderate values of Nymphaea nouchali and Potamogeton pectinatus (2.64% each) whereas, Nymphoides indica(1.76%), Lemna minor (1.53%) and Typha latifolia (1.12%) occurred in lower values. Among ferns, monolete (1.89%) and trilete spores(4.16%) are found in low to high value respectively.
Among arboreals, tree taxa include Terminalia bellirica and Dillenia pentagyna (1.06% each) in lower values, however, rest of the deciduous tree taxa like Shorea robusta, Syzygium cumini, Lagerstroemia parviflora, Adina cordifolia, Emblica officinalis, Salmalia malabaricum, Lannea coromandelica, Semecarpus anacardium, Meliaceae, Sapotaceae, Acacia catechu, Albizia lebbeck and Arecaceae are feebly represented along with scarce presence of semi evergreen tree taxa like Ilex sulcata, Artocarpus chaplasha, Schima wallichii and Symplocos racemosa. Among prominent shrubs, Melastoma malabathricum is dominant with a moderate value of2.25% followed by Ziziphus mauritiana (1.46%) and Oleaceae (1.10%) in lower values. Highland taxa including Pinus, Betula and Alnus are represented in improved frequency of 6.71%. Fungal remains are encountered in relatively lower values as compared to preceding two cases.
Pollen rain data from spider webs
Studying pollen and spores retrieved from the spider webs are of immense significance to understand the aerospora for meaningful comparison with the data on the differential pollen dispersal and deposition in a particular forest/unforested region, generated through other conventional methods viz., pollen catchers, moss cushions, etc. A total of seven spider web samples were procured, two each from within forest, forest edge and three from open-land area from closely placed branches of trees to understand the behaviour of air borne pollen grain in the light of pollen dispersal and deposition in and around the reserve forest and its implication in past vegetation and climate studies[27].
Within forest: The study of two samples has reflected comparatively higher values of Non Pollen Palynomorph (fungal remains and varia) which dominate the spectra and comprises of Nigrospora, Alternaria, Xylaria, Curvularia, Diplodia, Tetraploa, Helminthosporium, Cookeina and Microthyriaceous fruiting body as major fungal remains (32%) including insect wing, tracheids and epidermal peeling with stomata as varia. However, arboreal taxa include Syzygium cumini, Terminalia bellirica, Lagerstroemia parviflora, Artocarpus chaplasha, Bauhinia purpurea, Semecarpus anacardium, Dipterocarpus macrocarpus and Alstonia scholaris (30%), grasses (20%). Terrestrial herbs are composed of Chenopodiaceae, Amaranthaceae, Tubuliflorae, Evolvulus sp., Celosia argentea, Xanthium strumarium, Solanum sp. and Justicia simplex within 12%. Highland taxa (6%) are encountered as Pinus and Alnus.
Forest edge: The study of two samples has, predicted slight increment in the values of NPP dominating the spectra with fungal remains as Curvularia, Helminthosporium, Cookeina, Tetraploa, Xylaria and Diplodia (35%) and varia under 7%. Arboreal taxa are represented by Salmalia malabaricum, Syzygium cumini, Cesalpinia pulcherrima, Dillenia pentagyna, Emblica officinalis, Alstoniascholaris, Ziziphus mauritiana and Oleaceae (25%) along with grasses (16%) and open-land herb (9%) which mainly includes Lamiaceae, Brassicaceae, Acanthaceae, Verbenaceae, Tubuliflorae, Amaranthaceae, Mimosa pudica, etc. However, highland taxa like Pinus and Alnus show improved values upto 8% as compared to the preceding case.
Open-land: The study of three samples predict the dominance of NPP with fungal remains as Helminthosporium, Cookeina, Alternaria and germinating spore within 30% and varia (5%). Thearboreal taxa in the spectra show significantly reduced values from open-land spider meshes which are mainly represented by Alstonia scholaris, Terminalia bellirica, Syzygium cumini, Ziziphus mauritianaand Oleaceae within 12%. Grasses are improved by 30%, followed by open-land herbs (15%) including Impatiens balsamina, Celosia argentea, Xanthium strumarium, Rumex nepalensis, Polygonum plebeium, Plantago erosa, Brassicaceae, Acanthaceae, Euphorbiaceae, Apiaceae, Papaveraceae, Tubuliflorae, Liguliflorae and Artemisia parviflora. Highland taxa (8%) improved as compared to preceding spectra from within forest and forest edge.
Discussion
Palynological investigation have been conducted on 20 surface samples including 14 subsurface soil and six moss cushion procured from within forest across the open-land area of Panchratna RF ofGoalpara District. The study of pollen rain on subsurface soil and moss cushion has predicted the predominance of non-arboreals (49-65%) over arboreals (20-36%). The total pollen assemblage indicateexistence of semi-factual forest composition of tree elements like Shorea robusta Terminalia bellirica, Emblica officinalis, Syzygium cumini, Dillenia pentagyna, Lagerstroemia parviflora (as main ingredient) associated with Adina cordifolia, Lannea coromandelica, Semecarpus anacardium, Sapotaceae, Acacia catechu, Alstonia scholaris and Salmalia malabaricum as major deciduous forest taxa along with semi evergreen tree taxa like Ilex sulcata, Symplocos racemosa, Schima wallichii and Artocarpus chaplasha. However, Melastoma malabathricum, Ziziphus mauritiana and Oleaceae are incorporated as major prominent shrubby taxa in the assemblage.
Among non-arboreals, terrestrial herb such as Tubuliflorae, Liguliflorae, Artemisia, Xanthium, Caryophyllaceae, Amaranthaceae, Brassicaceae, Lamiaceae, Mimosa pudica, Acanthaceae and Plantagoare represented at moderately lower values. Tubuliflorae with average 6.05% pollen has comparatively good value, amply depicting the intense pastoral activities in the area of investigation as members of this family escape grazing because of its unpalatable nature to cattle and goats [28]. The special feature of this study indicate, underrepresentation of Shorea robusta, Terminalia bellirica, Lagerstroemia parviflora, Emblica officinalis, Dillenia pentagyna and Syzygium cumini as a dominant ingredients of the forest could be attributed due to poor preservation of their pollen in sediment despite their high pollen productivity [29]. However, the erratic representation of other arboreals could be inferred to low pollen productivity owing to prevalence of entomophily (e.g. Melia azedarach and Salmalia malabaricum) [30]. High pH value of soil as well as the microbial and chemical degradation of the pollen grains of taxa in the sub-surface soil might have been detrimental factors for the scarcity of pollen of some plants [31].
In sequel a set of seven spider webs were undertaken in this paper to evaluate air borne pollen within forest as well as open-land area successfully. The old spider meshes has yielded better pollen data in terms of quality and quantity. Interestingly, arboreal taxa represented in the meshes within forest and its outskirt mostly are of entomophilous as evidenced by Syzygium, Salmalia, Dillenia,Caesalpinia, Anacardiaceae, etc., comprising almost same values (25-30%) at the margin and core of the forest. Whereas, these values dwindles away (12%) when examined from open-land site. Ground vegetation is honestly portrayed consisting of grasses and terrestrial herbs in almost constant values in the forest area (25-32%) with inclination in the values of same (45%) from open-land. Higher valuesof fungal remains mostly grass pathogens are significant. Adequate Cerealia and Brassicaceae are proved as most apparently trapped pollen which remains suspended in the air for long or depending upon the exposure of the webs to the flowing air currents or wind. Highland taxa although in almost all the samples support long distance thermophilic pollen transport from upper reach. Judging all the substrates, it has been found that like moss and subsurface soil, spider webs also portray almost same type of face value picture in a forested area especially in regard to arboreals and grasses. Thus, it is concluded from the above observation that spider webs can also be taken as an additional tool for pollen analysis to understand the face value evaluation of extant vegetation in a forested region. Theexamination could also be helpful in evaluating the allergenicity of different pollen grains/spores in the area of investigation, causing asthma, hay fever, dermatitis and other disorders [32]. Allergic diseases can be controlled and symptoms can be minimized if we know what triggers those [33]. Thus, the study may be useful for allergologists too in establishing a right diagnosis and ultimately enable an improved quality of life for the inhabitants of the area of investigation [34]. The results of the study may be used in public awareness programmes about the health hazards caused by pollen grains.
Grasses (both Poaceae under < 60μm and Cerealia under > 60 μm) played the dominant role representing good frequencies in almost all samples as major ingredient of the ground vegetation (Table 3). Poaceae (grass) pollen have been categorized on the basis of their pollen sizes i.e., < 60μm (mostly wild type) and >60μm (Cerealia) for convenience in pollen interpretation. The occurrence of more Cerealia represents the acceleration in pastoral activity and wild types in general as vast open grassland. However, same may occur at patches within the forest may be attributable to other anthropic factors. Among ferns, monolete spores show higher values as compare to trilete spores in almost all the samples procured from the vicinity of woods, however, some monolete ferns are not of local nature which requires further investigation. Highland taxa (extraregional) have marked low frequencies in surface samples of forest and outskirts (2-4%) in contrast to open-land soil and spider webs (6-8%), attributed to free wind velocity in open areas. Fungal elements are mostly comprised of grass pathogens belonging to Ascomycetes and Deuteromycetes which are regularly represented in the sediment at reasonably good values. The plentiful record of the fungal spores, viz., Glomus, Microthyriaceae, Pleospora and Nigrospora, in particular, together with sporadic presence of Curvularia, Cookeina and Diplodia implies the availability of rich decaying organic matter on the forest floor for their propagation and dissemination [35]. Judging from the pollen assemblage it is evident that the set up of pollen distribution pattern has enabled to deduce the pollen/vegetation relationship within the forest and also provide a background data for deciphering past vegetation and climate change.
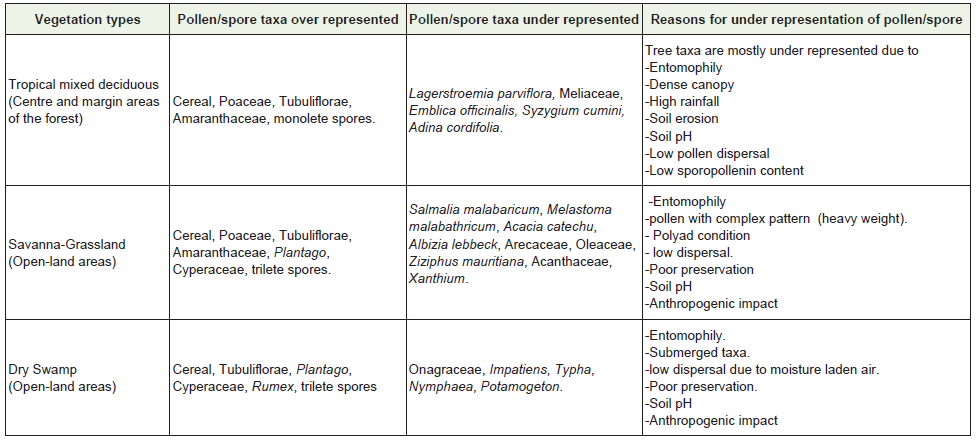
Table 3: Plant assemblage in respective vegetation type in the light of present pollen/spore data.
Conclusions
The dearth of recent pollen depositional data from Assam is a significant impediment towards establishing the Holocene vegetation history of this globally important, north-eastern region of India [36]. The relationships between current vegetation, the available environmental data and modern pollen are examined from Goalpara district for both natural and human-induced vegetation types, as an aid for palaeoenvironmental reconstructions. The palynodata produced from 20 surface pollen samples reflect the mixed deciduous forest comprising Terminalia, Lagerstroemia, Shorea, Emblica, Syzygium, Salmalia and Acacia under warm and humid climate in response to rainfall dissimilitude in the region. The pollen spectra have shown the dominance of non-arboreals (especially grass pollen and Asteraceae) and relatively lower frequencies of arboreals (APs; trees and shrubs). So, the pollen assemblages are not fully comparable to the actual set-up of vegetation in the area of investigation, so far as APs are concerned as some of the taxa are over-represented and some under-represented, which could be due to differences in pollen production, dispersal and preservation of taxa, depending on the plant species and climatic conditions. The minimal representation of core tree taxa in the palynoassemblage procured from the woods may be attributed to varied biotic and abiotic factors like entomophily; pollen with complex pattern (heavy weight); polyad condition; low pollen production; dispersal and preservation; soilpH and anthropogenic impact. Urban and agricultural land uses have interrupted the continuity of lower Assam forests and created forest islands. Pollen concentration and pH values are inversely proportional to each other, i.e. the concentration of pollen taxa decreases with the increase of pH values [37]. Thus, high pH values of the soil and oxidation coupled with erosion by fungi (and/or bacteria) could have been the other causes behind the fallacious display and recovery of pollen taxa in the samples [38,39]. However, a partial coherence between contemporary vegetation and surface pollen/spore data was observed (relative increase and decrease of arboreals in forest and open-land area respectively) which will be a potential base line data for the reconstruction of vegetation and climate change since late Quaternary.
Acknowledgements
We are highly grateful to Prof. Sunil Bajpai, Director, BSIP, Lucknow for providing necessary facilities and motivation for conducting the work. Thanks are also due to Dr. S.K. Basumatary,Scientist ‘B’, BSIP and forest officials in Assam for their kind help.
References
- Bradley RS (1985) Quaternary Palaeoclimatology: Methods of Palaeoclimatic Reconstruction. Boston, Allen & Unwin, London.
- Ranhotra PS, Bhattacharyya A (2013) Modern vegetational distribution and pollen dispersal study within Gangotri glacier valley, Garhwal Himalaya. Journal of the Geological Society of India 82: 133-142.
- Chauhan MS (1994) Modern pollen/vegetation relationship in the tropical deciduous sal forest in Madhya Pradesh. Journal of Palynology 30: 165-175.
- Singh G, Chapra SK, Singh AB (1973) Pollen rain from the vegetation of North-West India. New Phytology 72: 191-203.
- Champion HG, Seth SK (1968) A Revised Survey of Forest Types of India. New Delhi..
- Chauhan MS, Quamar MF (2013) Pollen rain deposition pattern in tropical deciduous Sal (Shorea robusta Gaertn.) forest in Shahdol District, Madhya Pradesh, India. Palaeobotanist 62: 47-53.
- Swati Dixit, Bera SK (2011) Mid-Holocene Vegetation and Climatic variability in Tropical deciduous Sal (Shorea robusta) forest of Lower Brahmaputra valley, Assam, Northeast India. Journal of the Geological society of India 77: 419-432.
- Bera SK (2000) Modern pollen deposition in Mikir hills, Assam. Palaeobotanist 49: 325-328.
- Basumatary SK, Dixit S, Bera SK, Mehrotra RC (2013) Modern pollen assemblages of surface samples from Cherrapunjee and its adjoining areas, Meghalaya, northeast India. Quaternary International 298: 68-79.
- Basumatary SK, Bera SK, Sangma SN, Marak G (2014) Modern pollen deposition in relation to vegetation and climate of Balpakram valley, Meghalaya, Northeast India: Implications for Indo-Burma palaeoecological contexts. Quaternary International 325: 30-40.
- Barboni D, Bonnefille R (2001) Precipitation signal in modern pollen rain from tropical forests, South India. Rev Palaeobot Palynol 114: 239-258.
- Anupama K, Ramesh BR, Bonnefille R (2000) Modern pollen rain from the Biligirirangan-Melagiri hills of southern Eastern Ghats, India. Rev Palaeobot Palynol 108: 175-196.
- Kutzbach JE (1976) The nature of climate and climatic variations. Quaternary Research 6: 471-480.
- Wilson CL & Matthews WH (eds) (1971) Inadvertent climate modification report of the study of man’s impact on climate (SMIC). Cambridge, Mass.: MIT Press.
- Mittermeier RA, Gil RP, Hoffmann M, Pilgrim J, Brooks T, et al. (2004) Hotspots revisited: Earth’s biologically richest and most endangered terrestrial ecosystems. Cemex, Mexico.
- Hazarika AK (2011) Over exploitation of forest resources its impact on the Environment. International Refereed Journal 26: 50-51.
- Gupta A (2009) Development of Barak valley: the Question of Sustainability In: Bhattacharjee, J.B. (Ed.) - Development strategies of Barak valley (Assam). Akansha Publishing House, New Delhi: 202-218.
- Swati Dixit, Bera SK (2012a) Holocene climatic fluctuation from lower Brahmaputra floodplain of Assam, Northeast India. Journal of Earth System Science 121: 135-147.
- Swati Dixit, Bera SK (2012b) Pollen rain studies in wetland environ of Assam, Northeast India, to interpret present and past vegetation. Journal of Earth Sciences and Engineering 5: 739-747.
- Bera SK (2003) Early Holocene pollen data from Mikir Hills, Assam, India. Palaeobotanist 52: 121-126.
- Karunakaran C (1974) Geology and mineral resources of the states of India. Part IV-Arunachal Pradesh, Assam, Manipur, Meghalaya, Mizoram, Nagaland and Tripura. Geological Survey of India Miscellaneous Publication 30: 1-24.
- Bera SK, Basumatary SK, Dixit S (2009) Studies on pollen morphology and phenological characteristics of some economically important arborescent taxa of Tropical forest, lower Brahmaputra Valley, Assam, North East India. J Palynol 43: 1-19.
- Bera SK, Trivedi A, Sharma C (2002) Trapped pollen and spores from spider webs of Lucknow environs. Current science 83: 1580-1585.
- Erdtman G (1943) An introduction to pollen analysis. Waltham, Mass. U.S.A.
- Nayar TS (1990) Pollen flora of Maharashtra State, India. Today and Tomorrow’s Printers & Publishers, New Delhi.
- Chauhan MS & Bera SK (1990) Pollen morphology of some important plants of tropical deciduous sal (Shorea robusta) forests, district Sidhi, Madhya Pradesh. Geophytology 20: 30-36.
- Singh S, Kar R, Khandelwal A (2010) Impact of modern pollen rain studies from south and Little Andaman Islands, India, to interpret present and past vegetation. Current Science 99: 1251-1256.
- Mazier F, Gallop D, Brun C & Butler A (2006) Modern pollen assemblage from grazed vegetation in western Pyrenes, France: a numerical tool for more precise reconstruction of past cultural landscapes. Holocene 16: 91-113.
- Quamar MF, Bera SK (2014) Surface pollen and its relationship with modern vegetation in tropical deciduous forests of southwestern Madhya Pradesh, India: a review. Palynology 38: 147-161.
- Quamar MF, Chauhan MS (2013) Modern pollen assemblages from teak (Tectona grandis Linn. F.) dominated tropical deciduous forest in southwestern Madhya Pradesh, India. The Palaeobotanist 62: 29-37.
- Sharma C (1985) Recent pollen spectra from Garhwal Himalaya. Geophytology 13: 87-97.
- Quamar MF, Chauhan MS (2011) Pollen analysis of spider webs from Khedla village, Betul District, Madhya Pradesh. Current science 101: 1586-1592.
- Guvensen A, Uysal I, Celik A, Ozturk M (2005) Analysis of airborne pollen falls in Canakkale, Turkey. Pak J Bot 37: 507-518.
- Altungolu MK, Toraman E, Temil M, Bicakci A, Kargioglu M (2010) Analysis of airborne pollen grains in Konya, Turkey. Pak J Bot 42: 765-774.
- Bera SK, Dixit Swati, Basumatary SK, Gogoi R (2008) Evidence of biological degradation in sediments of Deepor Beel Ramsar site, Assam as inferred by degraded palynomorphs and fungal remains. Current Science 95: 178-181.
- Bera SK, Dixit S (2010) Pollen analysis of Late Holocene lacustrine sediment from Jeypore reseve forest, Dibrugarh, Assam. Geological processes and Climate change. Edited by Dhruv Sen Singh and NL Chhabra: 85-94.
- Dimbleby GW (1957) Pollen analysis of terrestrial soils. New Phytologist 56: 12-28.
- Gupta HP, Yadav RR (1992) Interplay between pollen rain and vegetation of Tarai-Bhabar area in Kumaon Division, U.P., India. Geophytology 21: 183-189.
- Gupta HP, Sharma C (1985) Pollen analysis of modern sediments from Khasi and Jaintia hills, Meghalaya, India. Journal of Palynology 21: 167-173.