The Physio-Chemical Responses of Camellia Plants to Abiotic Stresses
Mainaak Mukhopadhyay1*and Tapan Kumar Mondal2
Corresponding author: Mainaak Mukhopadhyay, Department of Botany, University of Kalyani, Kalyani-741235, West Bengal, India,; E-mail: mainaakmu333@gmail.com
1Department of Botany, University of Kalyani, Kalyani-741235, West Bengal, India
2Division of Genomic Resource, National Bureau of Plant Genetic Resources, Pusa, New Delhi-110012, India
Citation: Mukhopadhyay M, Mondal TK. The Physio-Chemical Responses of Camellia Plants to Abiotic Stresses. J Plant Sci Res. 2014;1(1): 105.
Copyright © 2014 Mainaak Mukhopadhyay et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Plant Science & Research | ISSN: 2349-2805 | Volume: 1, Issue: 1
Submission: 18/02/2014; Accepted: 28/03/2014; Published: 02/04/2014
Abstract
Camellia species, being a woody perennial with more than 100 years of life span, it experiences several abiotic stresses throughout its life. Conventional breeding is protracted and restricted principally to selection, which escorts to taper down of its genetic base. Predominantly being strict monoculture cultivation, the plants countenance pest populations that are dreadful and unique. Young leavesare economically important parts and abiotic stresses are extremely detrimental to production. For instance, drought alone accounts for 40% loss of yield of tea per annum. Despite constraints, commendable efforts have been perpetrated to appreciate the physiological as well as biochemical alterations of an assortment of abiotic stresses encountered by these plants. Thus, this review presents a consolidatedaccount of the accomplishment and inadequacy of these tools and techniques hitherto applied to the plants. Expectedly, this will form a foundation for making further advances intended for improvement of tea and other economically important wild relatives, in particular, belongs to Camellia sp. To the best of our understanding, this is the first comprehensive compilation of such study in Camellia specieswith particular reference to tea [Camellia sinensis L. (O) Kuntze].
Keywords: Abiotic stress; Camellia; Physiological parameters; Biochemical analysis.
Introduction
Tea [Camellia sinensis (L.) O. Kuntze], of family Theaceae and an evergreen shrub, is indigenous to Indo-China region and is cultivated in humid and sub-humid tropical, sub-tropical, and temperate regions of the world, in about 52 countries [1], which grow mainly on acidic soils [2]. Apart from tea, a number of its wild species bear enormous significance. For instance, C. japonica, a wild species of tea, is cultivated due to its excellent floricultural splendor. Quite a few other wild species such as C. reticulata, C. sasanqua, and C. saluensis are also popular owing to their ornamental value [3]. Species such as C. oleifera, C. semiserrata, and C. chekiangolomy produce oil from mature seeds destined to pharmaceutical industry, albeit to a limited scale.
Stress evaluation and exploration of alleviatory actions are the two most active and kinetic research fields of tea science. In this study, progress in the research of abiotic stresses of tea plants have been appraised.
Abiotic Stress
Abiotic stresses are responsible for at least 65% diminution of teayield [4,5]. Being perennial, tea plants experience manifold abiotic stresses adversely influencing growth and yield, and therefore they need to tackle an array of multi-pronged stresses at some unspecified junctures of life cycle. To facilitate adaptation and survival, they have evolved counteractive mechanisms to endure stresses. Nevertheless, stresses beyond tolerance levels will unavoidably culminate in oxidative damage attributable to intensive production of reactive oxygen species (ROS) [6]. Oxidative stress is a phenomenon surfaces as a consequence of unevenness concerning the ROS production and competence of antioxidative defensive devices [7] [Figure 1].
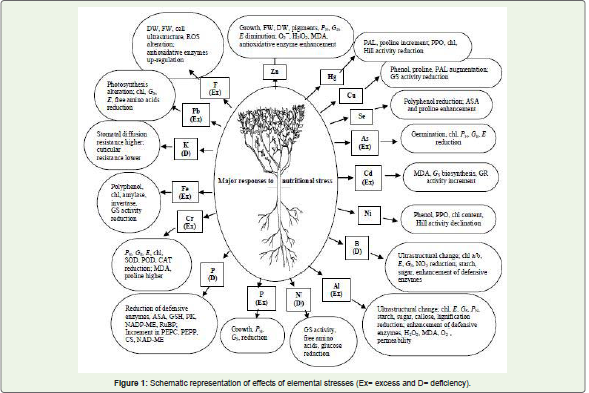
Figure 1: Schematic representation of effects of elemental stresses (Ex= excess and D= deficiency).
Macronutrients
The perennial plantation crops such as tea encounter typical predicament of element deficiency due to extended life span, unrelenting harvest without proper replenishment of nutrients and deterioration of soil fertility owing to human interference. Apart from deficiency, heavy metal contaminations due to constant handling and industrial emission have become a critical alarm to the tea plantationworldwide. Enormous improvements have been made till date in unveiling a handful of elements that result in an exponential amount of crop loss.
Nitrogen (N)
N is an essential macro element for plants. Tea plants require immense quantity of N not only for amino acid biosynthesis, but also for yield of secondary metabolites. Tea is low pH tolerant and favors ammonium (NH4+) because of quicker absorption. But N, as nitrate, acted as a stressor and reduced glutamine synthetase (GS) activity, N content, free amino acids, glucose and leaves became yellow under deficiency [8]. Likewise, in coffee plants, the rate of superoxide (O2-) formation increased under sunlight indicating increased photoprotective capacity per amount of photons absorbed. However, scorbate peroxidase (APX) activity reduced with hydrogen peroxide (H2O2) accumulation. Unsaturation of chloroplast membranes was higher in N deficient coffee plants [9]. The generation of ROS such as; O2-, H2O2 and hydroxyl radical (Oh•) damaged cellular components, protein, and membrane lipids. Plants responded to ROS by increased defensive enzymes pro duction under N deficiency suggesting the role of N as stress inducer. A positive correlation between peroxidase (POD) activity and tol erance to N deficiency affected water relations of C. sinensis by increasing stomatal resistance and transpiration rate (E) [10]. Similarly, Picea sitchensis, N deficiency reduced chlorophyll (chl) and carotenoid (car), photosynthetic rate (PN) and stomatal conductance (GS), and increased intercellular partial pressure of CO2(Ci) [11]. Similarly, under N deficiency, Fraxinus seedlings decreased uptake of N and phosphate, reduced leaf mass ratio and biomass [12] and conversely, increased N concentration, reduced chl, soluble protein and ribulose bi phosphate carboxylase oxygenase (Rubisco) of red pine trees drastically [13] indicating the probable role of N as stressor in excess also. however, problems with excess N are not very frequent in the natural ecosystems; it is widespread in areas with agricultural malpractices.
Phosphorus (P)
Due to a constituent of DNA, RNA, ATP and phospholipids, P is essential for plants. Tea is cultivated mainly on acidic soils where P deficiency is ubiquitous [14] and P deficiency led to lower activities of superoxide dismutase (SOD), APX, glutathione reductase (GR), catalase (CAT), monodehydroascorbate reductase (MDAR), dehydroascorbate reductase (DHAR) together with ascorbic acid (ASA) and reduced glutathione (GSH). But malondialdehyde (MDA) levels indicating active participation of antioxidant system against photo-oxidative injury [15]. P induced generation of singlet oxygen (1O2) and other ROS, exerted several harmful effects. Therefore, decrease in ASA, probably exposed P deficient plants to oxidative damages that compounded by decreased antioxidative enzyme activity. Conversely, P deficient treatments altered internal use efficiency. P deficiency, in tea, inhibited growth due to reduction of PN and GS that caused photoinhibition and photodamage of photosynthetic apparatus [16], and induced malate and citrate release accompanied by increased activity of PEP carboxylase, PEP phosphatase, citrate synthase (CS) and NAD-malic enzyme (NAD-ME) in addition to decreased pyruvate kinase (PK), NADP-malic enzyme (NADP-ME) and NADP-isocitrate dehydrogenase activities in roots. In the tea leaves, NADP-ME, NAD-ME and PK activities enhanced [17]. In Fraxinus sp., P deficiency, reduced 5-aminolevulinic acid biosynthesis and porphobilinogen synthase activity, N and P content, that in turn, decreased maximum quantum efficiency of PS-II, electron transport rate and PN [18]. In tea, P deficiency decreased the nutrients, and root Mg content. Leaves showed lower N and higher C and C/N ratios under deficiency [14]. P deficiency reduced ATP content terminating in a smaller amount of RuBP regeneration and CO2 assimilation. Energy dissipation increased to facilitate protection of P-deficient tea plants [14]. Excess light can be fatal for plants due to harmful ROS generation in the photochemical reaction centers. This happens when energy absorption exceeds the rate of carbon fixation. Hence, in order to survive, plants have evolved several protective processes by which energy dissipation has improved. Therefore, decrease in ASA and other antioxidative enzymes are liable to predispose plants to oxidative damages under P stress.
Iron (Fe)
Fe is an important macroelement of plants. Fe toxicity, in tea, declined chl and polyphenols along with decreased amylase, invertase, aspartate aminotransferase, glutamate synthase activity [19]. Polyphenols, the major alkaloids, decreased under Fe toxicity suggesting the gradual loss of protection to overcome oxidative damage and declined quality of tea [20]. In shade trees severe Fe deficiency resulted in chlorosis, reduced leaf as well as shoot growth leading to dieback of twigs [21]. Fe deficiency, in Quercus, also decreased chl and car contents, but decrease of chl was more. Fe deficiency caused 8% decrease in the dark-adapted efficiency of PS-II and 43% decrease in PS-II efficiency at steady-state photosynthesis [22].
Potassium (K)
K is one of the major macro elements that play a key role in a variety of movements of individual cells in certain tissues [23]. K deficient tea plants demonstrated increased stomatal diffusive resistance, reduced night opening and lower cuticular resistance [24]. In K-deficient Hibiscus, leaf water content, turgor potential, and PN, E, as well as GS were consistently lower, compared to K-sufficient plants [25]. Consistent lowering of gas exchange related parameters indicate that K deficiency may be fatal to plants because K+ is involved in guard cell movement.
Magnesium (Mg)
Mg is another crucial macroelement of plants and deficiency of Mg reduced glutathione and increased MDA in Citrus sinensis and C. grandis [26]. Hence, increase in MDA with deficient Mg is indicative of free radical generation. Mg toxicity induced development of coppery color all over the leaf surfaces and led to defoliations in tea plants, prevented Ca uptake because both are divalent cations with similar radius to each other. Synthesis and transport of amino acids in tea plants decreased under toxicity that also inhibited polyphenol synthesis of tea. Activities of nitrate reductase, glutamine synthetase, aspartate and alanine aminotransferase decreased under toxicity [27]. Mg deficiency, in Citrus reticulate, decreased leaf CO2 assimilation, carbohydrate contents, chl, car, CAT and Rubisco activity and impaired photosynthetic electron transport chain [28] indicating itspivotal role as a central molecule of chl.
Micronutrients
Trace elements are required in very little quantity but their importance for the growth of the plant is no way less than the major elements. Due to deficiency of these minor elements leaves, branches and fruits may not grow properly and they may even affect quality of products as well as production. These elements help in synthesis of hormones, enzymes, chl and absorption.
Boron (B)
B is an essential mineral nutrient for plants. Under starvation, NO3 concentrations diminished while NO2 increased in the young tea leaves and APX and POD activity increased in leaf and roots respectively [29]. Tea plantlets reduced growth, gas exchange parameters, pigments, dry weight (DW) and fresh weight (FW). Total and reducing sugar, starch, ASA and phenolics enhanced, but protein contents declined. Quantity of O2-, MDA, electrolyte leakage and H2O2 increased under deficiency. B deficiency up-regulated activities of POD, SOD and APX [30]; a similar observation was made in mulberry [31] which indicated that B deficiency provoked oxidative stress and the enzymes associated with the antioxidant system may be considered as stress markers [30]. Oxidative metabolism, the prime defense mechanism, brought about a cascade of biochemical transformations and implied the essence of B in tea. Besides, closing of stomata, increased abscisic acid (ABA) and proline indicated moisture stress under B deficiency [30]. Likewise, B deficiency increased phenolic compounds in leaves of olive plants [32]. Transmission electron micrograph (TEM) of cell walls elucidated thickened middle lamellae of leaf cells; a key deficiency marker because of its nonexistence in B-adequate cells. Under deficiency, the B-rhamnogalacturonan-II complex influenced the solidification of the middle lamella [30]. Likewise, due to B deficiency, phenol metabolites accumulated in olive leaves [32] and free amino acids increased in Lupinus [33].
Zinc (Zn)
Zn is a microelement, deficiency of which affects plants. Apart from other reasons, increase of P in the soil due to excessive phosphate applications induced Zn deficiency [34]. Zn stress (deficiency or toxicity) led to inhibition of growth, and FW and DW. Zn stress reduced PN, E and GS in conjunction with chl a and b [35]. Excess Zn, in vivo, altered photosynthetic parameters in Populus x euramericana [36]. O2-, MDA, h2O2 and electrolyte leakage elevated in deficient and toxic plants whereas total and reducing sugar and starch increased in optimum plants. Phenol content and of antioxidative enzyme activities intensified in Zn deficient plants indicating inflated ROS production under Zn stress [35]. Similarly, activities of APX, CAT, POD and SOD were increased in Zn-deficient and -sufficient mulberry [37] indicating their protective role. In sycamore maples, small quantities of Zn set off oxidative stress in older leaves because Zn reached sensitive sites inside leaf blade. They accelerated cell senescence resulting in chlorosis and necrosis [38]. Typical toxicity symptoms of Zn included leaf browning. UPASI-9 cultivars, under toxicity of 2000 mg kg-1 died within 15 d, while 1000 mg kg-1 took the toll by 36 d. Zn toxicity antagonized P, Mg and Fe absorption [39]. In Zn-scarce tea, youngest leaves turned narrow, strap-like, erect and formed rosette. Leaf blades became yellow and severely affected leaves became stunted with inward-curling margins; apical growth and branch extension retarded [34]. Chloroplast ultrastructures transformed under Zn excess and chloroplast membrane ruptured, thylakoid collapsed, adversely influencing photosynthesis. Similarly, mitochondria swelled and cristae disintegrated implying the failure of antioxidants to scavenge surplus ROS [35].
Aluminum (Al)
Al phytotoxicity is well-known on acidic soils in temperate and tropical regions. However, tea plants are Al hyper-accumulator and accumulate huge quantity in mature leaves [40] which is toxic for other crops. Al uptake by roots reduced the concentration of Ca, Mg and P. Al led to Mg deficiency in Norway and red spruce, Ca deficiency in loblolly pine and P deficiency in American and European beech [41].
In tea, Al at toxic concentration induced oxidative damage and Al contents increased in mature leaves. Pigments, PN, E and GS decreased but activated oxygen metabolism was evidenced by increasing MDA, O2-, H2O2 and membrane injury. Phenol, total sugar and starch decreased under toxicity but APX, GR, POD, CAT and SOD activities increased with elevated Al. Citrate and malate production also increased to minimize toxicity [20]. Besides, increased callose formation was a frequent response under toxicity [42]. TEM analysis of Al treated leaves, revealed significant membrane damage at 0.53 mM, due to loss of equilibrium between ROS formation and detoxification [43]. Capacity of Al accumulation depends on genotype because two Chinese cultivars, Pingyangtezao and Wuniuzao, performed differently as indicated by differential accumulation of chl and defensive enzyme activities [44]. Al and F, interaction had noteworthy effects on physiochemical parameters of tea because, Al alone enhanced root systems and SOD activity, but electrolyte leakage, POD vis-a-vis CAT activities declined; whereas, F increased enzyme activity but decreased permeability. Proline, POD, CAT and SOD activities remained lower under high Al: F, but when F increased, enzymes escalated, signifying injury [45]. Higher Al concentration drastically inhibited root elongation in Picea abies seedlings [46].
Al contents generally increased from young to mature leaf and the 6th mature leaf divulged that Al progressively increased from the centre towards the periphery [20]. Earlier studies revealed that, Al was detoxified by exclusion mechanism forming Al-ligand complexes and oxalate was a key chelating compound in tea roots [47] and oxalate exudation along with small quantity of malate, citrate and oxalate metabolisms were coupled to Al accumulation in roots [48]. Poly-Laspartate and poly-L-glutamate protected growth of pollen tube, in tea plants, from toxicity of Al by either interfering with Ca2+ transport or binding to calmodulin [49]. Cell suspension cultures of Picea rubens, to Al toxicity resulted in a loss of cell viability, inhibition of growth and a significant decrease in mitochondrial activity along with increased soluble protein [50]. Al reduced water permeability of red oak root cortex, and diffusion resistance of Picea and Abies. Reduction of stomatal aperture was observed in many species because Al altered membrane permeability, stomatal aperture and root surface area. Alinduced shoot damage in peach and coffee. In coffee shoots, toxicity symptoms included chlorosis and necrosis of younger leaves and spotty chlorosis of older leaves [41].
Chromium (Cr)
Cr is a widespread natural contaminant and its accumulation in soil creates a disparaging effect on tea plants. It not merely affects production and quality, in addition jeopardizes human health through food chain. Cr toxicity decreased root and shoot length and protein, chl, car, proline and nitrogenase in Pongamia glabra, Cassia auriculata, Pithecellobium dulce, Azadirachta indica and Tamarindus indicus [51] and induced metabolic modifications, alterations in photosynthesis in Genipa americana [52].
Cr diminished chl content, PN, GS and E along with the number of new buds, and weight of tea [53]. Increasing Cr toxicity lowered photosynthesis indicators, along with altered antioxidation and modified resistance. Activities of defensive enzymes like, SOD, POD and CAT diminished, while proline, MDA and conductivity increased in C. sinensis [54]. Probably Cr increased free radical generation, as indicated by increased MDA production. The content of MDA, a peroxidation product of membrane lipids, is often used as an indicator of oxidative damage. Thus, pigment and protein destruction indicate inefficacy of antioxidants to scavenge excessive ROS generated because of Cr toxicity.
Lead (Pb)
Pb is a well-known pollutant having the ability to function as a stressor. Pb substantially affected seed germination, seedling growth and DW in Thespesia populnea [55]. In Pb contaminated soil, phytotoxic symptoms appeared in tea plants along with reduction in biomass, caffeine and free amino acid but catechin content increased and the roots accumulated higher quantity of Pb [56]. Pb toxicity impaired photosynthesis and decreased chl, PN, GS and E significantly [53]. Nevertheless, tolerance to Pb toxicity depends upon the genotypes of tea. Longjing43 showed an enhancement in PN when both Longjing43 and Zhenong117 were subjected to Pb toxicity. POD and SOD activities increased under toxicity in Zhenong117 but in longjing43, POD activity boosted up initially while SOD activity increased gradually [57]. Besides, Pb inhibited seed germination of Pinus helipensis [58]. Pb toxicity increased the growth of Longjing43 (a tea variety), but reduced in Zhenong117. Conversely, at a lower level, Pb decelerated the net PN of Zhenong117, but enhanced that of Longjing43. In Zhenong117, POD and SOD activities increased with increasing Pb whereas in Longjing43 POD activity of increasedinitially and then decreased [59].
Fluorine (F)
F is phytotoxic as it can alter a series of metabolic pathways. Amygdalis communis seedlings, under increasing F, decreased chl, Ca, Mg, and starch and sugar content of leaves. Mineral concentration, mainly Mn, showed a major decrease at 2.5 mM F [60]. Tea plants accumulated large quantity of F innately, but under increasing Ca, the uptake was affected possibly due to the effect of Ca on the properties of cell wall or membrane permeability or altered F speciation [61]. Under high F stress (>0.32 mM), chloroplast membranes ruptured, thylakoid expanded and degenerated and reduced PN [62]. F also mitigated the toxic effect of Al and stimulated pollen tube growth in tea [63]. In conifers, F and SO2 initially induced chlorosis on needle apices and brachyblasts that became greyish-brown due to the death of cells and gradually fell down. This defoliation process ensued rapidly in the presence of HF and gas mixtures that contained HF [64]. Small leafed Chinese tea variety accrued very high F compared to their large leafed equivalent while the Assam variety contained the lowest F [65]. Toxic F concentrations decreased FW, DW, chl and PN whereas CAT and guaiacol peroxidase (GPX) activities increased considerably along with H2O2, MDA and proline [62]. Increased H2O2 in the F toxic leaves probably triggered Fenton/Haber-Weiss reaction associated with generation of OH• radical and lipid peroxidation. However, increased activities of antioxidative enzymes minimized stress to an extent beyond which growth retarded.
Mercury (Hg)
Hg pollution has become a vital concern due to its environmental impact on a universal scale. Germination of seeds of Platanus occidentalis, Pinus echinata, and Pinus taeda under mercuric nitrate and methyl mercury chloride solutions were decreased, with P. echinata being the least sensitive. Germination was inhibited more when seeds were exposed to methyl mercury chloride than the other. Organic species of mercury was found to be more toxic than inorganic counterparts [66]. Hg can also induce oxidative stress in tea. Upon exposure, Hg decreased phenol contents, PPO and chl along with Hill reaction, but, on the contrary, PAL activity and proline contents increased under Hg toxicity [67]. Tea plants exposed to high Hg reduced PN, growth and led to root browning as well as leaf chlorosis [68]. Thus, it may be recommended that antioxidative defence system did not adequately protect the tissues under severe Hg stress.
Copper (Cu)
Cu is an essential element for all forms of life, acting as the prosthetic group of many enzymes. However, Cu toxicity inhibited root elongation of both P. pinea and P. pinaster seedlings within 3 d of exposure. Root cortex thickened significantly and lignin synthesis increased in P. pinaster, and decreased cell elongationcaused by increased permeability and cell-wall lignification was the main mechanisms of Cu toxicity in Pinus [69]. Cu ions, in tea plants, oxidized thiol bonds causing disruption in protein structure and functions [68]. Cu stress increased phenol, proline and PAL activity in tea but PPO and chl reduced [67] whereas glutamine synthase activity decreased [70]. Chinary cultivars were more tolerant than Assamica, because the later accumulated more ROS, showed higher activity of phytochelatin synthase [68].
Excessive soil Cu hindered the extension of young branches of apple, decreased the content of active Fe and chl significantly, and declined the activity of CAT in young leaves [71].
Selenium (Se)
Although Se is a trace element, but toxic if present in excess. Prunus species showed reduced growth with incremental Se treatments. Se induced a partial stomatal closure, as evidenced by GS, resulting in a reduction in net assimilation, and thus a decrease in dry-mass production [72]. In green tea, vitamin C content remained lower and during storage decline was much higher in Se deficient plants. The color of green tea extract in Se-enriched green tea was more stable compared to the counterpart during the storage period. The sweetness and aroma of Se-enriched green tea were also significantly higher [73]. It also indicated that Se may play positive role in increasing the quality parameters.
Nickel (Ni)
Ni is an essential nutrient deficiency of which culminated in mouse ear disease in pecan [74] and biochemically disrupted the ureide pathway, the urea cycle, the citric acid cycle, and the shikimic acid pathway [75]. Pinus sylvestris, upon exposure to Ni, augmented higher sucrose accumulation, signifying interruptions in carbohydrate metabolism. Trees exposed to Ni had higher content of condensed tannins compared with controls [76]. Ni executed the role of a stressor in tea plants also, because it decreased phenols, PPO and chl content along with Hill activity [67] but proline contents and PAL activity were increased in tea [67]. PAL had widely been studied concerning its induction by various environmental factors such as light, wounding excision and infection that may be considered as stress generators. In plants, PAL is an important intermediate in the metabolic pathway to phenylpropanoid metabolism and thus performs defense-related functions [77]. In presence of excess Ni the activity of PAL as well as other stress markers increased indicating the potent role of Ni in inducing stress.
Arsenic (As) and Cadmium (Cd)
As concentrations elevated in soils due to geochemistry or human activities. As toxicity-decreased chl in Pistacia lentiscus and Tamarix gallica, but MDA increased considerably [78]. Enhanced As++ decreased germination and twig growth rigorously along with instantaneous reduction in chl, PN, GS and E [79]. In As and Cd polluted soil, feeder roots played the buffering role and minimum quantity of As was transported to the shoots. Cd persuaded growth, photosynthesis and chl synthesis when present in a tolerable quantity but surplus quantity impaired the plants [79] but P. tremula exhibited growth inhibition and changed gene expressions and a decreased level of proteins [80]. Similarly, Cd exposure decreased PN, E and biomass [81], chl and protein in tea, but enhanced MDA, expression of γ-glutamylcysteine synthetase, glutathione synthetase, and GR [82], which implied the defensive activity of these enzymes under excess Cd. In spruce needles, the soluble fractions of GPX initially increased and decreased subsequently. Increased cell wall bound POD activity was marked under extended treatment [83]. Effect of Cd on tea indicated that, chl remained higher in spring than summer and reduced with an increment in Cd, which enhanced MDA and soluble sugar in spring tea [84] indicating the role of Cd as a potent stressor. Cd concentration changed phenol metabolism in callus culture depending on the origin of the tissues. In leaf and stem originated callus, it decreased biomass and flavans but uniqueness remained unaffected in calli derived from roots. Simultaneously lignin content in the calli of root and stems increased, while remained unaltered in the leaf [85]. Appreciable inhibition of the nitrate reductase activity was found in Silene cucubalus [86]. Growth, APX and CAT activities inhibited along with H2O2 accumulation in response to Cd in poplar roots [87]. A significant reduction in plant height, biomass, stem diameter, leaf area and weight of leaves in hybrid poplar was observed under Cd toxicity. CAT activity and proline accumulation decreased significantly, but SOD activity reduced in roots and increased in the leaves under toxicity [88]. A gradual decrease in growth of Dalbergia sissoo with increasing Cd levels was reported [55].
Manganese (Mn)
Mn is an essential micronutrient required for the normal growth of higher plants and may become toxic when present in excess [89]. Mn deficiency depressed leaf photosynthetic capacity in plants of Carya illinoinensis, primarily by reducing the number of PSII units per unit area of leaf whereas Populus cathayana, under humid condition accumulated more Mn, decreasing growth, chl and activities of antioxidant enzymes than the dry climate population [90]. In Citrus, Mn-excess decreased CO2 assimilation, GS, increased intercellular CO2, whereas Rubisco activity decreased. Chl a fluorescence (O-J-I-P; chl fluorescence kineics) transients from Mn-excess leaves showed increased O- and decreased P-steps, accompanied by positive L- and K- bands, indicating Mn-excess leaves had increased damage of oxygen evolving complex and less energy exchange between independent PSII units. Mn-excess decreased light energy transformation efficiency (Fv/Fm) but increased relative variable fluorescence at I-steps (VI) and energy dissipation. These leaves displayed higher MDAR, GR, SOD, CAT and guaiacol peroxidase (GPX) activities and contents of antioxidants [91]. Thus, it indicated that Mn excess leaves were damaged by oxidative stress, which was possibly due to the overreduction of the photosystem as a result of slower dark reactions resulting from decreased Rubisco and other enzymes. Hence, it can be suggested that the defensive system of plant regulated the changes of enzyme activities to facilitate the defensive function against toxic Mn.
Humidity and Drought
Moisture stress is a major limitation for plant growth, and to counteract, several physiological and antioxidative mechanisms have evolved. Tea plants under drought or inundation declined water potential (ψ) and decelerated growth. Inherently, even tea seeds are desiccation sensitive (Mondal et al. 2001). Activities of PPO and POD decreased along with PN and metabolism, but conductivityincreased [92]. Tender leaves had high diffusion resistance whereas drought tolerant clones showed increased relative water content [93] but clones that minimized water loss, endured drought. Soil moisture deficiency severely affected PS-II reaction centre [94] but PN increased in irrigated tea compared to drought-affected [95]. Similarly, maximal fluorescence (Fm), variable fluorescence (Fv), Fv/Fm and activity potential of PS-II (Fv/Fo) decreased leading to non-photochemical quenching [96]. Moisture deficiency reduced shoot: root ratio, leaf area, stem diameter, net CO2 assimilation and rearranged dry matter partitioning to roots [97]. Soil water content influenced GS that affected E and played a key role in shaping water status of leaves. Decreased GS in response to increased irradiance, leaf temperature (TL) and air vapor pressure debit played a key role in photoinhibition of tea [98]. Similarly, in Corylus avellana L., GS progressively decreased with increased long-term water deficiency [99]. At TL, of 20-30°C in tea, PN decreased at a rate of 0.053 μmol m-2s-1°C-1. However, under water deficiency, optimum TL remained around 20-24°C while PN reduced significantly [100,95].Total chl content decreased under dehydration. High humidity generated maximum PN whereas low temperature along with low soil moisture eased PN. Under inundation, photosynthetic photon flux density (PPFD) decreased prominently [101]. Under drought, the amount of dry matters partitioned to leaf, stem and harvested shoots decreased by 80-95% [102]. Drought increased conductivity [103] in tea. Solute leakage is the indicator of stresses such as metal toxicity; drought etc that gives rise to increased ROS, which in turn, increases membrane permeability and culminates in electrolyte leakage.
Water deficiency led to increase in car, ABA, GSH, and GPX activities but reduced proline, H2O2 and O2-, however subsequent rehydration led to recovery [104]. A positive correlation documented that the activity of Rubisco, POD and PPO enzymes served as markers to monitor drought tolerance [103.1055]. Tea cultivars (TV-18, -26, T-78 and HV-39), under water stress, enhanced phenol and proline along with increased phenylalanineammonia lyase (PAL), but prolonged drought exhibited reverse trend. Drought augmented accumulation of proteins but protracted stress, diminished them in T-78 and HV- 39 [106]. Drought stress followed by rehydration treatments, reduced ASA and GSH in TV-1, 20, 29 and 30, and among them, TV-1 had the highest activities of GR and CAT [107] indicating the severe effect of drought. China cultivars, namely; Zhengong 113, Fudingdabai, Yunqi and Zisun under drought, increased proline. PN and E was the highest in Zhengong 113, and Zisun was the most affected with lowest PN and E [108]. Tea seeds under desiccation increased proline, H2O2 content, APX and SOD activities [109.110] indicating that the level of osmoprotectants increased. Before the appearance of visual symptoms of drought, dehydrin, ABA ripening protein, calmodulin-binding protein, GPX and cinnamoyl CoA reductase increased in tea leaves [111]. Similarly, under desiccation APX and SOD activities increased in tea seeds. Increased activity of CAT, ASA, and other antioxidants, scavenged ROS that altered redox status to become deleterious [109]. CaCl2 application thwarted drought induced oxidative damage by increasing DW, proline, phenol, activity of SOD, POD, CAT and GR enzymes and plummeted H2O2 and MDA [112]. Accumulation of K, Ca, Mn and B accelerated during drought recovery and they positively modulated enzymes like, SOD, CAT, POX, PPO and GR that scavenge ROS suggesting that drought caused oxidative damages to plants [113].
Moisture deficiency reduced PAL activity in UPASI-2, -8 and -9 clones and reduced the synthesis of EGCG and ECG due to molecular rearrangement with elevated leaf temperature. Similarly, flooding deteriorated tea quality, due to reduction of gallic acid and caffeine contents leading to reduced synthesis of epitheaflavicacid, epitheaflavic acid-3′-gallate and theaflavic acid [114]. Drought tolerant clones had been found to translocate more photosynthates to shoots under moisture stress [115]. Downpour for longer period led to lowering of EGC, epicatechin, ECG, EGCG but increased catechins. However, relative humidity significantly deteriorated EGC, TC and caffeine [116]. Thus, it seems that flooding as well as drought influences the quality of made tea because the prime components for the supremacy of tea i.e., the quality parameters were declined.
Continuous evapo-transpiration eventually became fatal. Hence, application of antitranspirants minimized the effect and ‘film type’ antitranspirants were more competent than ‘stomatal type’. Phenyl mercuric acetate (PMA) is useful due to its inefficient translocation and short life span [117]. Alongside ABA, commercial antitranspirants such as Raliidhan and Antistress are effective due to increased stomatal diffusion resistance, ψ, relative turgidity, and reduced E [118]. A linear relationship between total DW and the ratio between E and mean saturation vapor pressure deficit in clonal tea plants manifested marked significance [119].
Leaf necrosis and defoliation in Eucalyptus pilularis under severe drought stress [120] indicated the effect of drought. In apple, emissions of α-pinene, β-pinene and limonene were negatively correlated with rainfall which supported that drought resulted in higher formation of secondary metabolites [121].
Temperature
Temperature is an important abiotic stress, which below 13°C adversely affect various biosynthetic activities of tea [122]. Low temperature is one of the key stresses that tea bushes necessitate to cope with. They are also susceptible to frost damage that culminates in scorching of young pluckable shoots and defoliations [123]. During winter, low temperatures alone or in combination with low irradiation induced oxidative stress in plants. It had been found that genotypes having shorter dormancy period displayed elevated activities of defensive enzymes due to accrual of ROS. Efficient scavenging of toxic oxygen species led to lower accumulations during winter months coupled with reduced winter dormancy [124]. Photosynthesis canbe a potential marker to understand the effects of low temperature stress on plants. Low temperature tolerant tea genotypes were found to possess higher MDA, proline and car content along with enhanced ROS scavenging machinery than the susceptible clones [122]. Therefore Netto [122] recommended that clones, for instance; TTL-1, TTL-4 and UPASI-9 being low temperature and frost tolerant were more suitable compared to clones like, TTL-6, SM/OM/54 and SMP- 1 for a region having recurrent low temperature situations because plantation of these clones will ensure the production of augmented yield and quality [122]. Low temperature induced albino tea cultivar, reversed the albino phenomenon when the temperature was above 20°C, resulting sharp decline in amino acids content. In albino tea, the development of chl from etioplast and the accumulation of chl a and b continued to be the same at 15°C; thus culminating in albino shoots. Experimental evidences specified that chl content reduced with decrease in temperature and leaves were in need of mature chloroplasts at 15°C; rather, etioplasts developed [125]. Cold tolerance of grafted and non-grafted tea plants were correlated to PN in vivo [126]. Exposure of tea buds to proline and betaine decreased cold stress, enhanced methylglyoxal and MDA, and increased GR and glutathione-S-transferase activity [127]. Tea growing under the shade of Grevillea robusta had substantially lower transpiration rates than unshaded tea because shades reduced E by minimizing irradiance that strike the canopy and also by reducing canopy temperature [119]. PN and GS of Quercus alba decreased between 35 and 40°C, whereas in Q. rubra the parameters increased over this range [128], thus indicating vulnerability of white oak to temperature. Therefore, temperature stresses can be detrimental for growth and development of tea and other woody plants. Nevertheless, tea plants have evolved complex mechanisms to cope with such unfavorable conditions and thus increased levels of ROS that trigger changes in plants are scavenged by antioxidants up to a certain extent.
Ultraviolet-B (UV-B) Irradiation
UV-B is harmful to plants [129] as it reduced expression and synthesis of chl a/b-binding protein and the D1 polypeptide of PSII and inhibited growth [130,131]. Short-term irradiation of UV-B stimulated accumulation of total catechins. Conversely, excessive UV-B irradiation suppressed catechin accumulation. EGCG increased more rapidly than other catechins under irradiation [132]. UV-B impeded size of callus forming cells and promoted deposition of phenolic compounds on cell-wall and intercellular spaces [133]. UV-B induced chl-bearing cell formation and PS-II activity stimulated in the phototrophic cells of callus culture [134]. PN, E, and TL of tea leaves were affected by photosynthetic photon flux densities (PPFD) and experimental evidences revealed that PPFD of 2,200 μmol m-2 s-1 affected PN but E and TL of maintenance leaves linearly increased from 200-2,200 μmol m-2 s-1 PPFD [135]. B deficient tea plants mitigated the reduction of photosynthetic energy conversion under intermediate and highly illuminated circumstances. Under intermediate light intensity, CO2 assimilation was the highest and produced lower activity of APX, SOD and proline content [136].
Environmental factors acts upon the contents of tea plants. Darkness inhibited synthesis of flavanols, declined gallocatechin and leucoanthocyanin contents. The leucoanthocyanins content of darkened stems decreased much less than the leaves[137]. Leaves appreciably gained phenyl propanoids, benzenoids and phenylalanine under shade, whereas upstream metabolites like, shikimic acid, prephenic acid, and phenylpyruvic acid decreased [138].
Impact of Various Stresses on Wild Relatives of Tea
Camellia is the largest genus of the family Theaceae that includesmore than 325 species [139]. In a conservative estimation, there are more than 3,000 cultivated varieties of ornamental Camellia worldwide of which more than 2,500 have been registered in the American Camellia Society [140]. Camellia sp. has significantly different growth habits in different distribution areas due to diverse climatic conditions [141].
The relationship between the processing temperature and the cell injury rate showed that C. oleifera appeared as the best suitable species for high-temperature zones whereas C. vietnamensis suited for temperate regions [141]. High temperature increased soluble protein, sugar and free proline in C. oleifera leaves [142]. Conversely, under prolonged chilling, the leaves of C. rusticana showed no visible changes while C. japonica proliferated after 200 d. The decline in the rate of soluble carbohydrate content in C. rusticana remained about one-sixth of that in C. japonica. The PN of C. japonica decreased to half of its initial value after 140 d, while that of C. rusticana remained same even after roughly one year [143].
Light intensity, radiation etc also affected physiological and biochemical characteristics of wild relatives of Camellia. C. nitidissima changed both qualitative and quantitative characters under changed light environment. Relative light intensity that was lesser than 20%; correlated leaf length and width, but beyond more than 20%,insignificant correlations occurred. Low light intensity induced development of short-narrow leaves, while high light intensity stimulated longer and wider leaves [144]. Under shade, the stomata of C. rusticana closed constantly, however japonica, remained open even after 90 d [143]. γ radiation, increased SOD and POD activity in ‘Ruanzhi 2’ and ‘Ruanzhi 3’ upto 1000 rad but decreased under 3000 rad whereas ‘Ruanzhi 11’ and ‘Yue 15’ yielded the lowest under 3000 rad [145]. Type of planting and density of plants also influenced PN because low planting yielded higher PN than higher planting population [146].
In C. oleifera, Al, under acidic conditions increased chl a, b, GS and PN but decreased when Al concentration increased. Simultaneously PN also affected biomass accumulation [147]. C. oleifera required suitable concentrations of Al and P for optimal growth [148] but under Al toxicity and P inadequacy altered growth and photosynthesis rate. Lime in combination with P led to increase in PN, GS and decreased intercellular CO2 concentration. Simultaneously car/chl changed significantly, suggesting role of car in photo-protection [149]. Another study revealed that P deficiency led to higher activity of acid phosphatase in some varieties [150]. In fact, leaf age and leaf position had been reported to be two important factors, which influenced PN [151].
Leaf blight disease of C. oleifera is one of the major diseases, which causes premature defoliation. It had been found that Bacillus subtilis was able to control up to 84.7% of the diseases [152]. C. sasanqua pollen grain produced shorter pollen tubes due to growth inhibitory substances but when soaked in acetone or diethyl ether produced longer pollen tubes suggesting that acetone or diethyl ether probably played as growth stimulant [153]. Simulated acid rain produced inhibition of seed germination, increased MDA,but reduced POD, SOD, chl a, chl b, and their ratio (www. mt.china-papers.com; 2010). Spraying paclobutrazol on the leaves of C. oleifera seedling inhibited shoot growth but root growth was promoted, and chl content, PN and protein enhanced [154].
Dehydration also affected pivotal antioxidants because dehydrated seedlings increased the activity of PPO, POD and CAT activity along with MDA and relative conductivity [155]. Under 0% - 30% dehydration resistance value of leaf, decreased while the values of root and stem showed the same trend under 0% - 25% of dehydration [156].
Conclusion and Future challenges
Tea plants, being woody perennial, experience quite a lot of abiotic stresses. Although, conventional breeding is successful for varietal improvement of tea, yet, they are focused on to the improvement of yield, quality, drought and few diseases tolerance. While significant amount of achievement have been made in improving yield and quality parameters of tea, other attempts have marginally improved the objective. This is primarily due to the fact that most of the stresses are complex in nature, controlled by several genetic elements. Thus, prior to any attempt to progress further; it is essential to understand the physiological as well biochemical basis of such stresses, which later formulate the groundwork for studying the molecular basis. In tea, several studies have been accomplished which gives an apparent idea about the parameters involved.
Tea is a hyper-accumulator and accumulates Al in such an extent that is toxic to other plants. Thus identification of the molecule behind such tolerance will facilitate to understand the mechanism which in turn will assist to develop the Al tolerant plant in other species. In general, detection of the key biochemical molecules associated with physiological parameters would aid nursery selection foragronomically important traits and will assist molecular breeding ultimately. Approaches like association mapping can only be successful if morphological, physiological as well as biochemical scoring are successful. In tea where conventional breeding is limited, association mapping shows tremendous scope for varietal improvement for complex traits.
Acknowledgements
The authors are thankful to Department of Biotechnology, Govt of India and Tea Board, Ministry of Commerce for financial help to carry out part of the research. Authors wish to dedicate this article to Prof N. K. Jain, a noted tea scientist of international repute for his contribution toward the development of Indian Tea Industry.
References
- Mondal TK, Bhattacharya A, Laxmikumaran M, Ahuja PS (2004) Recent advances of tea (Camellia sinensis) biotechnology. Plant Cell Tiss Org Cult 76: 195-254.
- Lin ZH, Chen LS, Chen RB, Zhang FZ, Jiang HX, et al. (2009) CO2 assimilation, ribulose-1,5-bisphosphate carboxylase /oxygenase, carbohydrates and photosynthetic electron transport probed by the JIP-test, of tea leaves in response to phosphorus supply. BMC Plant Biol 9: 43.
- Mondal TK (2002) Camellia biotechnology: a bibliographic search. Int J Tea Sci 1: 28-37.
- Mondal TK (2003) Micropropagation of Tea (Camellia sinensis). Micropropagation of woody trees and fruits 75: 671-719.
- Waheed A, Hamid FS, Shah AH, Ahmad H, Khalid A, et al. (2012) Response of different tea (Camellia sinensis L.) clones against drought stress. J Master Environ Sci 3: 395-410.
- Smirnoff N (1993) The role of active oxygen in the response of plants to water deficit and desiccation. New Phytol 125: 27-58.
- Apel K, Hirt H (2004) Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annu Rev Plant Biol 55: 373-399.
- Ruan J, Gerendás J, Härdter R, Sattlemacher B (2007) Effect of nitrogen form and root-zone pH on growth and nitrogen uptake of tea (Camellia sinensis) plants. Ann Bot 99: 301-310.
- Pompelli MF, Martins SCV, Antunes WC, Chaves ARM, DaMatta FM (2010) Photosynthesis and photoprotection in coffee leaves is affected by nitrogen and light availabilities in winter conditions. J Plant Physiol 167: 1052-1060.
- Nagarajah S (1981) The effect of nitrogen on plant water relations in tea (Camellia sinensis). Physiol Plantarum 51: 304-308.
- Chandler JW, Dale JE (1995) Nitrogen deficiency and fertilization effects on needle growth and photosynthesis in Sitka spruce (Picea sitchensis). Tree Physiol 15: 813-817.
- Wu C, Wang Z, Fan Z, Sun H (2004) Effects of nitrogen stress on nutrient uptake by Fraxinus mandshurica seedlings and their biomass distribution. Ying Yong Sheng Tai Xue Bao 15: 2034-2038.
- Takeshi I, Tatsuro N (2003) Effects of high nitrogen load and ozone on forest tree species. Eurasian J For Res 6: 155-170.
- Lin ZH, Chen LS, Chen RB, Peng A (2009) Effects of phosphorus deficiency on nutrient absorption of young tea bushes. J Tea Sci 29: 295-300.
- Lin ZH, Chen LS, Chen RB, Zhang FZ (2012) Antioxidant system of tea (Camellia sinensis) leaves in response to phosphorus supply. Acta Physiol Plant 34: 2443-2448.
- Salehi SY, Hajiboland R (2008) A high internal phosphorus use efficiency in tea (Camellia sinensis L.) plants. Asian J Plant Sci 7: 30-36.
- Lin ZH, Chen LS, Chen RB, Zhang FZ, Jiang HX, et al. (2011) Root release and metabolism of organic acids in tea plants in response to phosphorus supply. J Plant Physiol 168: 644-652.
- Wu C, Fang Z, Wang Z (2004) Effect of phosphorus stress on chlorophyll biosynthesis, photosynthesis and biomass partitioning pattern of Fraxinus mandchurica seedlings. Ying Yong Sheng Tai Xue Bao 15: 935-940.
- Hemalatha K, Venkatesan S (2011) Impact of iron toxicity on certain enzymes and biochemical parameters of tea. Asian J Biotechnol 6: 384-394.
- Mukhopadyay M, Bantawa P, Das A, Sarkar B, Bera B, et al. (2012) Changes of growth, photosynthesis and alteration of leaf antioxidative defence system of tea (Camellia sinensis (L.) O. Kuntze) seedling under aluminum stress. Biometals 25: 1141-1154.
- Neely D (1976) Iron deficiency chlorosis of shade trees. Journal of Arboriculture 128-130.
- Gogorcena Y, Molias N, Larbi A, Abadia J, Abadia A (2001) Characterization of the responses of cork oak (Quercus suber) to iron deficiency. Tree Physiol 21: 1335-1340.
- Marschner H (1995) Mineral Nutrition of Higher Plants. 2nd Ed. Academic Press, New York.
- Nagarajah S (1979) The effect of potassium deficiency on stomatal and cuticular resistance in tea (Camellia sinensis). Physiol Plantarum 47: 91-94.
- Egilla JN, Davies Jr FT, Boutton TW (2005) Drought stress influences leaf water content, photosynthesis, and water-use efficiency of Hibiscus rosa-sinensis at three potassium concentrations. Photosynthetica 43: 135-140.
- Yang GH, Yang LT, Jiang HX, Li Y, Wang P, Chen LS (2012) Physiological impacts of magnesium-deficiency in Citrus seedlings: photosynthesis, antioxidant system and carbohydrates. Trees 26: 1237-1250.
- Venkatesan S, Jayaganesh S (2010) Characterisation of magnesium toxicity, its influence on amino acid synthesis pathway and biochemical parameters of tea. Res J Phytochem 4: 66-67.
- Tang N, Li Y, Chen L (2012) Magnesium deficiency-induced impairment of photosynthesis in leaves of fruiting Citrus reticulata trees accompanied by up-regulation of antioxidant metabolism to avoid photo-oxidative damage. J Plant Nutr Soil Sci 175: 784-793.
- Hajiboland R, Bastani S, Rad SB (2011) Photosynthesis, nitrogen metabolism and antioxidant defense system in B-deficient tea (Camellia sinensis (L.) O. Kuntze) plants. J Sciences 22: 311-320.
- Mukhopadhyay M, Ghosh PD, Mondal TK (2013) Effect of boron deficiency on photosynthesis and antioxidant responses of young tea (Camellia sinensis (L.) O. Kuntze) plantlets. Russ J Plant Physiol 60: 633-639.
- Tewari RK, Kumar P, Sharma PN (2010) Morphology and oxidative physiology of boron-deficient mulberry plants. Tree Physiol 30: 68-77.
- Liakopoulos G, Karabourniotis G (2005) Boron deficiency and concentrations and composition of phenolic compounds in Olea europaea leaves: a combined growth chamber and field study. Tree Physiol 25: 307-315.
- Alves M, Chicau P, Matias H, Passarinho J, Pinheiro C, Ricardo CP (2011) Metabolic analysis revealed altered amino acid profiles in Lupinus albus organs as a result of boron deficiency. Physiol Plant 142: 224-232.
- Nelson S (2006) Zinc deficiency in tea (Camellia sinensis). Plant Dis PD-34.
- Mukhopadhyay M, Das A, Subba P, Bantawa P, Sarkar B, et al. (2013) Structural, physiological and biochemical profiling of tea plantlets (Camellia sinensis (L.) O. Kuntze) under zinc stress. Biol Plantarum 57: 474-480.
- Di Baccio DR, Tognetti, Minnocci A, Sebastiani L (2009) Responses of the Populus × euramericana clone I-214 to excess zinc: Carbon assimilation, structural modifications, metal distribution and cellular localization. Environ Exp Bot 67:153-163.
- Tewari RK, Kumar P, Sharma PN (2008) Morphology and physiology of zinc-stressed mulberry plants. J Plant Nutr Soil Sci 171: 286-294.
- André O, Vollenweider P, Günthardt-Goerg MS (2006) Foliage response to heavy metal contamination in Sycamore Maple (Acer pseudoplatanus L.). For Snow Landsc Res 3: 275-288.
- Venkatesan S, Hemalatha KV, Jayaganesh S (2006) Zn toxicity and its influence on nutrient uptake in tea. Amer J Plant Physiol 1:185-192.
- Matsumoto H, Hirasawa E, Morimura S, Takahashi E (1976) Localization of aluminum in tea leaves. Plant Cell Physiol 17: 627-631.
- Schaedle M, Thornton FC, Raynal DJ, Tepper HB (1989) Response of tree seedlings to aluminum. Tree Physiol 5: 337-356.
- Lian C, Oiwake Y, Yokota H, Wang G, Konishi S (1998) Effect of aluminum on callose synthesis in root tips of tea (Camellia sinensis L.) plants. Soil Sci Plant Nutr 44: 695-700.
- Li C, Xu H, Xu J, Chun X, Ni D (2011) Effects of aluminium on ultrastructure and antioxidant activity in leaves of tea plant. Acta Physiol Plant 33: 973-978.
- Yu C, Pan Z, Chen J, Fan D, Wang X (2012) Effects of Al (3+) on growth and physiological characteristics of tea plant (Camellia sinensis). Plant Nutr Fertilizer Sci 2012-01.
- Wang X, Liu P, Luo H, Xie Z, Xu G, Yao J, Chen K (2009) Effect of Al and F interaction on physiological characteristics of tea plants. Acta Horticulturae Sinica 2009-09.
- Godbold DL, Fritz E, Huttermann A (1988) Aluminum toxicity and forest decline. Proc Natl Acad Sci USA 85: 3888-3892.
- Morita A, Yanagisawa O, Takatsu S, Maeda S, Hiradate S (2008) Mechanism for the detoxification of aluminum in roots of tea plant (Camellia sinensis (L.) Kuntze). Phytochemistry 69: 147-153.
- Morita A, Fujii Y, Yokota H (2001) Effect of aluminium on exudation of organic acid anions in tea plants. Plant Nutrition 92: 508-509.
- Konishi S, Ferguson IB, Putterill J (1988) Effect of acidic polypeptides on aluminium toxicity in tube growth of pollen from tea (Camellia sinensis L.). Plant Sci 56: 55-59.
- Minocha R, McQuattie C, Fagerberg W, Long S, Noh EW (2001) Effects of aluminum in red spruce (Picea rubens) cell cultures: Cell growth and viability, mitochondrial activity, ultrastructure and potential sites of intracellular aluminum accumulation. Physiol Plantarum 113: 486-498.
- Unnikannan P, Baskaran L, Chidambaram ALA, Sundaramoorthy P (2013) Chromium phytotoxicity in tree species and its role on phytoremediation. Insight Bot 3: 15-25.
- Barbosa RMT, Almeida AF, Mielke MS, Loguercio LL, Mangabeira PAO, et al. (2007) A physiological analysis of Genipa americana L.: A potential phytoremediator tree for chromium polluted watersheds. Environ Exp Bot 61: 264-271.
- Tang Q, Li X, Zhu X, Huang Y, Yang H (2008) Effects of plumbum and chromium stress on the growth of tea plants. Southwest J Agric Sci 21: 156-162.
- Tang J, Xu J, Wu Y, Li Y, Tang Q (2012) Effects of high concentration of chromium stress on physiological and biochemical characters and accumulation of chromium in tea plant (Camellia sinensis L.). Afr J Biotechnol 11: 2248-2255.
- Kabir M, Iqbal MZ, Shafiq M, Farooqi ZR (2008) Reduction in germination and seedling growth of Thespesia populnea L., caused by lead and cadmium treatments. Pak J Bot 40: 2419-2426.
- Wu Y, Liang Q, Tang Q (2011) Effect of Pb on growth, accumulation and quality component of tea plant. Procedia Eng 18: 214-219.
- Han WY, Shi YZ, Ma LF, Ruan JY, Zhao FJ (2007) Effect of liming and seasonal variation on lead concentration of tea plant (Camellia sinensis (L.) O. Kuntze). Chemosphere 66: 84-90.
- Nakos G (1979) Lead pollution: fate of lead in soil and its effects on Pinus haplenis. Plant Soil 50: 159-161.
- Luo Y, Kang M, Ren M (2004) Effect of lead pollution on tea plant growth and activity of its protective enzymes. J tea 2004-04.
- Elloumi N, Abdallah FB, Mezghani I, Rhouma A, Boukhris M (2005) Effect of fluoride on almond seedlings in culture solution. Fluoride 38: 193-198.
- Ruan J, Ma L, Shi Y, Han W (2004) The impact of pH and calcium on the uptake of fluoride by tea plants (Camellia sinensis L.). Ann Bot 93: 97-105.
- Li C, Zheng Y, Zhou J, Xu J, Ni D (2011) Changes of leaf antioxidant system, photosynthesis and ultrastructure in tea plant under the stress of fluorine. Biol Plantarum 55: 563-566.
- Konishi S, Miyamoto S (1983) Alleviation of aluminum stress and stimulation of tea pollen tube growth by fluorine. Plant Cell Physiol 24: 857-862.
- Rozhkov AS, Mikhailova TA (1993) The effect of fluorine containing emissions on conifers. pp ix + 142.
- Fung KF, Zhang ZQ, Wong JWC, Wong MH (2003) Aluminum and fluoride concentrations of three tea varieties growing at Lantau island, Hong Kong. Environ Geochem Health 25: 219-232.
- Jean-Philippe SR, Labbé N, Damay J, Franklin JA, Hughes K (2012) Effect of mercuric compounds on pine and sycamore germination and early survival. AJPS 3: 150-158.
- Basak M, Sharma M, Chakraborty U (2001) Biochemical responses of Camellia sinensis (L.) O. Kuntze to heavy metal stress. J Environ Biol 22: 37-41.
- Yadav SK, Mohanpuria P (2009) Responses of Camellia sinensis cultivars to Cu and Al stress. Biol Plantarum 53: 737-740.
- Arduini I, Godbold DL, Onnis A (1995) Influence of copper on root growth and morphology of Pinus pinea L. and Pinus pinaster Ait. seedlings. Tree Physiol 15: 411-415.
- Rana NK, Mohanpuria P, Yadav SK (2008) Expression of tea cytosolic glutamine synthetase is tissue specific and induced by cadmium and salt stress. Biol Plantarum 52: 361-364.
- Liu CS, Shi YX, Ma L, Ye YL, Yang JH, Zhang FS (2000) Effect of excessive copper on growth and metabolism of apple trees. Acta Metall Sin 6: 451.
- Pezzarossa B, Remorini D, Piccotino D, Malagoli M, Massai R (2009) Effects of selenate addition on selenium accumulation and plant growth of two Prunus rootstock genotypes. J Plant Nutr Soil Sci 172: 261-269.
- Huang Y, Xu J, Hu Q (2005) Effect of selenium on preservation quality of green tea during autumn tea-processing season. J Agric Food Chem 53: 7444-7447.
- Wood BW, Reilly CC, Nyczepir AP (2004) Mouse ear of Pecan: A nickel deficiency. Hort Sci 39: 1238-1242.
- Bai C, Reilly CC, Wood BW (2006) Nickel deficiency disrupts metabolism of ureides, amino acids, and organic acids of young pecan foliage. Plant Physiol 140: 433-443.
- Roitto M, Rautio P, Julkunen-Tiitto R, Kukkola E, Huttunen S (2005) Changes in the concentrations of phenolics and photosynthates in Scots pine (Pinus sylvestris L.) seedlings exposed to nickel and copper. Environ Pollut 137: 603-609.
- MacDonald MJ, D’Cunha (2007) A modern view of phenylanine ammonia lyase. Biochem Cell Biol 85: 273-282.
- Moreno-Jime´nez E, Esteban E, Carpena-Ruiz RO, Pen˜alosa JM (2009) Arsenic-and mercury-induced phytotoxicity in the Mediterranean shrubs Pistacia lentiscus and Tamarix gallica grown in hydroponic culture. Ecotoxicol Environ Safety 72: 1781-1790.
- Tang Q, Zhu X, Li X, Tan H (2008) Effects of As and Cd stress on the growth of tea plants. J Henan Agric Sci 2008-03.
- Kieffer P, Dommes J, Hoffman L, Hausman JF, Renaut J (2008) Quantitative changes in protein expression of cadmium-exposed poplar plants. Proteomics 8: 2514-2530.
- Shi Y, Ruan J, Ma L, Han W, Wang F (2008) Accumulation and distribution of arsenic and cadmium by tea plants. J Zhejiang Univ Sci B 9: 265-270.
- Mohanpuria P, Rana NK, Yadav SK (2007) Cadmium induced oxidative stress influence on glutathione metabolic genes of Camellia sinensis (L.) O. Kuntze. Environ Toxicol 22: 368-374.
- Radotic K, Ducic T, Mutavdžić D (2000) Changes in peroxidase activity and isoenzymes in spruce needles after exposure to different concentrations of cadmium. Env Exp Bot 44: 105-113.
- Xia J, Lan H (2008) Effects of cadmium stress on growth of tea plant and physiological index in leaves of mengshan tea. J Tea Sci 56-61.
- Zagoskina NV, Goncharuk EA, Alyavina AK (2007) Effect of cadmium on the phenolic compounds formation in the callus cultures derived from various organs of the tea plant. Russ J Plant Physiol 54: 237-243.
- Mathys W (1975) Enzymes of heavy metal-resistant and non-resistant populations of Silene cucubalus and their interactions with some heavy metals in vitro and in vivo. Physiol Plant 33:161-165.
- Schutzendubel A, Polle A (2002) Cadmium and H2O2 induced oxidative stress in Populus x canescens roots. Plant Physiol Biochem 40: 577-584.
- Nikolic N, Kojic D, Pilpovic A, Pajevic S, Krstic B, et al. (2008) Responses of hybrid poplar to cadmium stress: Photosynthetic characteristics, cadmium and proline accumulation, and antioxidant enzyme activity. Acta Biologica Crasoviensia 50: 96-103.
- 89.Mukhopadhyay M, Sharma A (1991) Manganese in cell metabolism of higher plants. Bot Rev 57: 117-149.
- Millaleo R, Reyes-DÃaz M, Ivanov AG, Mora ML, Alberdi M (2010) Manganese as toxic element for plants: Transport, accumulation and resistance mechanisms. J Soil Sci Plant Nutr 10: 470-481.
- Li Q, Chen LS, Jiang HX, Tang N, Yang LT, et al. (2010) Effects of manganese-excess on CO2 assimilation, ribulose-1,5-bisphosphate carboxylase/oxygenase, carbohydrates and photosynthetic electron transport of leaves, and antioxidant systems of leaves and roots in Citrus grandis seedlings. BMC Plant Biol 10: 42.
- Mondal TK, Bhattachraya A, Sharma M, Ahuja PS (2001) Induction of in vivo somatic embryogenesis in tea (Camellia sinensis) cotyledons. Current Science 81: 297-300.
- Sandanam S, Gee GW, Mapa RB (1981) Leaf water diffusion resistance in clonal tea (Camellia sinensis L.): Effects of water stress, leaf age and clones. Ann Bot 47: 339-349.
- Wijeratne MA, Formham R, Anandacumaraswamy A (1998) Water relations of clonal tea (Camellia sinensis L.) with reference to drought resistance: II. Effect of water stress. Tropical Agric Res Ext 1:74-80.
- Smith BG, Burgess PJ, Carr MKV (1994) Effects of clone and irrigation on the stomatal conductance and photosynthetic rate of tea (Camellia sinensis). Exp Agric 30: 1-16.
- Guo CF, Sun Y, Tang YH, Zhang MQ (2009) Effect of water stresson chlorophyll fluorescence in leaves of tea plant. Chin J Eco-Agric 17: 560-564.
- Bore JK (2008) Physiological responses of grafted tea (Camellia sinensis L.) to water stress. JKUAT abstracts of postgraduate thesia. Accessed on 3rd March.
- De Costa WAJM, Mohotti AJ, Wijeratne MA (2007) Ecophysiology of tea. Braz J Plant Physiol 19: 299–-332.
- Schulze ED, Küppers M (1979) Short-term and long-term effects of plant water deficits on stomatal response to humidity in Corylus avellana L. Planta 146: 319-326.
- Smith BG, Stephens W, Burgess PJ, Carr MKV (1993) Effects of light, temperature, irrigation and fertilizer on photosynthetic rate in tea (Camellia sinensis). Exp Agric 29: 291–306.
- Hajra NG, Kumar R (2002) Responses of young tea clones to subtropical climate: Effects on photosynthetic and biochemical characteristics. J Plant Biol 29: 257–264.
- Burgess PJ, Carr MKV (1996) Responses of young tea (Camellia sinensis) clones to drought and temperature. II. Dry matter production and partitioning. Experimental Agric 32: 377–394.
- Yang Y, Zhuang X, Hu H (1987) Effect of the soil water content on the physiological process of tea plant. J Tea Sci 7: 23–28.
- Upadhyaya H, Panda SK (2004) Responses of Camellia sinensis to drought and rehydration. Biol Plantarum 48: 597–600.
- Jeyaramraja PR, Jayakumar D, Pius PK, Kumar RR (2002) Application of Rubisco, peroxidase and polyphenol oxidase as markers for productivity and drought tolerance in tea. J Plant Biol 29: 315–320.
- Chakraborty U, Dutta S, Chakraborty BN (2002) Response of tea plants to water stress. Biol Plantarum 45: 557–562.
- Upadhyaya H, Panda SK, Dutta BK (2008) Variation of physiological and antioxidative responses in tea cultivars subjected to elevated water stress followed by rehydration recovery. Acta Physiol Plantarum 30: 457–468.
- Wu B, Pan G (1995) Studies on physiological and biochemical response to water stress in tea plant. Acta Agricultarae Universitatis Chekianensis 21:451–456.
- Chen Q, Yang L, Ahmad P, Wan X, Hu X (2011) Proteomic profiling and redox status alteration of recalcitrant tea (Camellia sinensis) seed in response to desiccation. Planta 233: 583–592.
- Hajiboland R, Bastani S (2012) Tolerance to water stress in boron deficient tea (Camellia sinensis) plants. Folia Horticulturae 24: 41-51.
- Gupta S, Bharalee R, Bhorali P, Bandyopadhyay T, Gohain B, et al. (2012) Identification of drought tolerant progenies in tea by gene expression analysis. Funct Integr Genomics 12: 543–563.
- Upadhyaya H, Panda SK, Dutta BK (2011) CaCl2 improves post-drought recovery potential in Camellia sinensis (L) O. Kuntze. Plant Cell Rep 30: 495–503.
- Upadhyaya H, Dutta BK, Sahoo L, Panda SK (2012) Comparative effect of Ca, K, Mn and B on post-drought stress recovery in tea [Camellia sinensis (L.) O Kuntze]. Am J Plant Sci 3:443–460.
- Jeyaramraja PR, Pius PK, Kumar RR, Jayakumar D (2003) Soil moisture stress-induced alterations in bioconstituents determining tea quality. J Sci Food Agric 83: 1187–1191.
- Jeyaramraja PR, Raj Kumar R, Pius PK, Thomas J (2003b) Photoassimilatory and photorespiratory behavior of certain drought tolerant and susceptible tea clones. Photosynthetica 41: 579–582.
- Wang LY, Wei K, Jiang YW, Cheng H, Zhou J, et al. (2011) Seasonal climate effects on flavanols and purine alkaloids of tea (Camellia sinensis L.). Eur Food Res Technol 233: 1049–1055.
- Nagarajah S, Ratnasooriya GB (1977) Studies with antitranspirants on tea (Camellia sinensis L.). Plant Soil 48: 185–197.
- Handique AC, Manivel L (1990) Effect of certain antitranspirants in tea. Two Bud 37:20–23.
- Anandacoomaraswamy A, De Costa WAJM, Shyamalie HW, Campbell GS (2000) Factors controlling transpiration of mature field-grown tea and its relationship with yield. Agr Forest Meteorol 103: 375–386.
- Weggler K, Carney C, Stone C (2008) Effect of fertiliser and water supply on the growth, nutrient status and photochemical efficiency of Eucalyptus pilularis seedlings in a phosphorus-deficient soil. Australian Forestry 71: 54–63.
- Vallat A, Gu H, Dorn S (2005) How rainfall, relative humidity and temperature influence volatile emissions from apple trees in situ. Phytochemistry 66: 1540–1550.
- Netto LA, Jayaram KM, Haridas P, Puthur JT (2005) Characterization of photosynthetic events and associated changes in various clones of tea (Camellia sinensis L.) under low temperature conditions. J Plant Biol 48: 326-331.
- Mondal TK (2003) Frost management of Tea. The Assam Review and Tea News. 92: 8-12.
- Vyas D, Kumar S, Ahuja PS (2007) Tea (Camellia sinensis) clones with shorter periods of winter dormancy exhibit lower accumulation of reactive oxygen species. Tree Physiol 27: 1253–1259.
- Du YY, Chen H, Zhong WL, Wu LY, Ye JH, et al. (2008) Effect of temperature on accumulation of chlorophylls and leaf ultrastructure of low temperature induced albino tea plant. Afr J Biotechnol 7: 1881–1885.
- Luo Y-P, Wu S, Kang M-L (2002) Photosynthetic characteristic and cold resistance of two-year old grafted tea plant in winter. J Zhejiang Univ Agric Life Sci 28: 397–400.
- Kumar V, Yadav SK (2009) Proline and betaine provide protection to antioxidant and methylglyoxal detoxification systems during cold stress in Camellia sinensis (L.) O. Kuntze. Acta Physiol Plant 31: 261–269.
- Sharkey TD, Singsaas EL, Vanderveer PJ, Geron C (1996) Field measurements of isoprene emission from trees in response to temperature and light. Tree Physiol 16: 649–654.
- Frederick JE, Snell HE, Haywood EK (1989) Solar ultraviolet radiation at the Earth’s surface. Photochem Photobiol 50: 443–450.
- Stapleton AE (1992) Ultraviolet radiation and plants: Burning questions. Plant Cell 4: 1352–1358.
- A-H-Mackerness S, Jordan BR, Thomas B (1997) UV-B effects on the expression of genes encoding proteins involved in photosynthesis. In: Lumsden PJ (editors) Plant and UV-B: Responses to environmental change. Cambridge University Press, Cambridge, UK, 113–114.
- Zheng XQ, Jin J, Chen H, Du YY, Ye JH, et al. (2008) Effect of ultraviolet B irradiation on accumulation of catechins in tea (Camellia sinensis (L) O. Kuntze). Afr J Biotechnol 7: 3283–3287.
- Zagoskina NV, Dubravina GA, Alyavina AK, Goncharuk EA (2003) Effect of ultraviolet (UV-B) radiation on the formation and localization of phenolic compounds in tea plants callus cultures. Russ J Plant Physiol 50: 270–275.
- Zagoskina NV, Alyavina AK, Gladyshko TO, Lapshin PV, Egorova EA, Bukhov NG (2005) Ultraviolet rays promote development of photosystem-II photochemical activity and accumulation of phenolic compounds in the tea callus culture (Camellia sinensis). Russ J Plant Physiol 52:731–739.
- Barman TS, Baruah U, Saikia JK (2008) Irradiation influences tea leaf (Camellia sinensis L.) photosynthesis and transpiration. Photosynthetica 46: 618–621.
- Hajiboland R, Bastani S, Rad SB (2011) Effect of light intensity on photosynthesis and antioxidant defense in boron deficient tea plants. Acta Biol Szeged 55: 265–272.
- Forrest GI (1969) Effects of light and darkness on polyphenol distribution in the tea plant (Camellia sinensis L.). Biochem J 113: 773–781.
- Yang Z, Kobayashi E, Katsuno T, Asanuma T, Fujimori T, et al. (2012) Characterisation of volatile and non-volatile metabolites in etiolated leaves of tea (Camellia sinensis) plants in the dark. Food Chem 135: 2268–2276.
- Mondal TK (2011) Wild Crop Relatives: Genomics and Breeding Resources Planation and Ornamental Crops, editor: C. Kole, Springer, USA.
- Mondal TK, Bhattacharya A, Laxmikumaran M, Ahuja PS (2004) Recent advances of tea (Camellia sinensis) biotechnology. Plant Cell Tiss Org Cult 76:195–254.
- He XY, Ye H, Ma JL, Zhang RQ, Chen GC, et al. (2012) Semi-lethal high temperature and heat tolerance of eight Camellia species. ΦYTON 81:177–180.
- Wang G, Chen L, Kou L, Yang Y, Feng F, et al. (2012) Effects of high temperature stress on osmotic adjustment substances of 25 varieties of Camellia oleifera. J Henan Agric Sci 2012–04.
- Kume A, Tanaka C, Matsumoto S, Ino Y (1998) Physiological tolerance of Camellia rusticana leaves to heavy snowfall environments: The effects of prolonged snow cover on evergreen leaves. Ecological Res 13: 117–124.
- Hu XH, Li JW, Jiang QS, Zhao RF (2010) Responses of leaf characters of Camellia nitidissima to different light environments. Guihaia 2010–03.
- Huang Y, Yuan Z, Zhong W, Zhang H, Dong B (2009) Effects of ~ (60)Co-γ radiation on isozyme activity of Camellia Oleifera seedling. Nonwood Forest Res 2009–02.
- He Y, Lu F (2008) Study on the photosynthesis of the different density of Camellia Oleifera. Modern Agric Sci 2008–03.
- He G, Liu Q, Peng S (2010) Effect of Aluminum toxicity on photosynthetic characters of wild Camellia oleifera under acidic conditions. Hubei Agric Sci 2010–07.
- Tan X, Yuan J, Li Z, Ye S, Jiang Z (2011) Effects of aluminum and phosphate on material distribution and roots characteristics and activity of Camellia oleifera seedlings. J Central South Univ Forestry Tech 2011–12.
- He G, Zhang J, Hu X, Wu J (2011) Effect of aluminum toxicity and phosphorus deficiency on the growth and photosynthesis of oil tea (Camellia oleifera Abel.) seedlings in acidic red soils. Acta Physiol Plantarum 33:1285–1292.
- Chen L, Chen Y, Wang R, Ma L, Peng S, et al. (2011) The effects of phosphorus deficiency stress on activities of acid phosphatase in different clones of Camellia oleifera Abel. Chinese Agric Sci Bull 2011–31.
- Rui Y, Yong-Zhong C (2009) Influencing factors on photosynthetic characteristic of superior clones of Camellia oleifera-leaf age and leaf position. Chinese Agric Sci Bull 25: 113-118.
- Li He, Zhou G, Zhang H, Song G, Liu J (2011c) Study on isolated pathogen of leaf blight and screening antagonistic bacteria from healthy leaves of Camellia oleifera. Afr J Agric Res 6: 4560–4566.
- Iwanami Y (1973) Acceleration of the growth of Camellia sasanqua pollen by soaking in organic solvent. Plant Physiol 52: 508–509.
- Hu Z, Shen W, Zhang Y (1993) Physiological effect of paclobutrazol on the growth of Camellia oleifera seedling. J Fujinan College Forestry 1993–02.
- Cao Z, Hu J, Shu Q, Wen J, Zhan W, Xu G, Bu J (2011) Effects of water stress on physical characteristics and survival rate of container seedling in Camellia oleifera. Nonwood Forest Res 2011–04.
- Cao Z, Zhang S, Liu C, Hu J, Zhang X, Shu Q (2011a) Effects of dehydration degree on the physiological and chemical characteristics of Camellia oleifera seedlings. J Anhui Agric Univ 2011–03.