Research Article
Specific Single Chain Antibodies (Scfvs)Against Influenza A Virus Using Phage Display Technique
Samaneh Alizadeh1, Foroogh Nejatollahi1,2*, Afagh Moattari3 and Bahareh Moazen1
1Recombinant Antibody Laboratory, Department of Immunology, Shiraz University of Medical Sciences, Shiraz, Iran
2Shiraz HIV/AIDS Research Center, Shiraz University of Medical Sciences, Shiraz, Iran
Corresponding author: Foroogh Nejatollahi, Associate Professor, Department of Immunology, Shiraz University ofMedical Sciences, Iran; E-mail: nejatollaf@sums.ac.ir
Citation: Alizadeh S, Nejatollahi F, Moattari A, Moazen B. Specific Single Chain Antibodies (Scfvs) Against Influenza a Virus Using Phage DisplayTechnique. J Immunol Vaccine Technol. 2014;1(1): 102.
Copyright © 2014 Foroogh Nejatollahi et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Immunology and Vaccine Technology | Volume: 1, Issue: 1
02/07/2014; Accepted: 26/08/2014; Published: 28/08/2014
Reviewed & Approved by Dr. Om Parkash, Scientist,Immunology Laboratory, National JALMA Institute forLeprosy and Other Mycobacterial Diseases, India
Abstract
Introduction: 2009 H1N1 swine-origin Influenza A virus with human pandemic potential emerged in 2009 and caused a number of mortality. Two highlyconserved regions 173-181 and 227-239 has recognized in Hemagglutinin of H1N1 Influenza virus. Immunity to the different subtypes of Influenza virus couldobtain with high affinity antibodies against conserved epitopes of the virus. In the present study, a phage antibody display library of scFv was applied for theselection of scFv antibodies against the conserved sequences of the hemagglutinin.
Materials and Methods: Panning process was used to isolate specific scFvs against highly conserved neutralizing epitopes of Hemagglutinin. PCRand fingerprinting were done on the selected clones. Phage ELISA was performed to evaluate the specificity of the clones selected against the epitopes.
Results: Two common patterns gainst each peptide were obtained which represented the isolation of two specific scFvs, scFv-I and svFv-II with thefrequencies of 25% (5/20) and 55% (11/20) against peptide 1 and scFv-I’ and scFv- II’ with the frequencies of 20% (4/20) and 75% (15/20) against peptide2. Phage ELISA showed the reactivity of the scFv antibodies with the related epitopes while no reaction with unrelated epitopes and other negative controlswere obtained.
Conclusion: Results demonstrated the isolation of two specific scFvs against two conserved neutralizing sequences of Hemagglutinin of Influenzavirus A. Due to several advantages of scFv, these antibodies are introduced as valuable agents in clinics. Results suggest further evaluation of the specificscFvs against Influenza A virus for their use in clinical applications.
Keywords: Rabies virus; Growth Kinetics; Fluorescent Activated Cell Sorter (FACS) analysis; Cell-ELISA; FAT
Introduction
Influenza infection in humans is one of the most importantrespiratory diseases in all over the world. The infection could befetal specially in children [1]. Influenza virus A, is responsible forprevalence of pandemics and the most of the yearly flu epidemics[2]. This type is further categorized into the subtypes according tothe properties of Hemagglutinin (HA) and neuraminidase (NA)antigens on the surface of viral particle [3]. The unexpected variabilityof the Influenza A virus makes annual epidemics in population andis the principal reason of the lack of impressive prevention againstInfluenza infection to date [4].
In April of 2009, a swine-origin influenza H1N1 virus (S-OIV)has appeared. 2009 pandemic H1N1 influenza A virus is a reassortantstrain of the influenza virus containing gene segments from swine, avian and human [5]. 2009 pandemic H1N1 influenza A virus isantigenically similar to the 1918 pandemic H1N1 Influenza A virus[6]. Until 2013, pandemic H1N1 has led to more than 1.4 millioninfections with about 25,000 deaths worldwide [7]. The most of theserious consequences caused by influenza infection occurred inindividuals with underlying medical conditions. The highest mortalityrates were found in immunocompromised individuals, patients withchronic neurological disease and respiratory disease [8]. Pregnantwomen were also under threat [9]. Due to Ser31Asn substitutionin the transmembrane Matrix 2 protein, the 2009 pandemic H1N1Influenza A virus is resistant to Amantadine antiviral drug. 2009pandemic H1N1 Influenza A virus is sensitive to Oseltamivir andZanamivir antiviral drugs [10,11].
Hemagglutinin is the main antigen responsible for theattachment of virus to the specific receptors on the surface of host cells, providing entrance into the host cell via endocytosis. It definesthe virulence, host ranges and transitivity of influenza viruses. Theprecursor of hemagglutinin (HA0) is synthesized in the infected hostcells and undergoes a enzymatic cleavage to obtain two subunits,HA1 and HA2 [12]. HA exhibit conserved and variable domain[13]. Hu et al [14] reported seven conserved epitopes of HA1 of 2009pandemic H1N1 Influenza A virus containing 5-23, 63-81, 142-158,178-195, 201-216, 246-269, and 277-287, which are responsible forattachment of the virus to the swine cells. According to the previousresearches conducted by Kubota-Koketsu et al [15] and Yamashita[16], two highly conserved epitopes 173-181 and 227-239 werereported which antibodies against these epitopes could neutralizethe different subtypes of influenza virus. This glycoprotein also hasa principal role in the emergence of human pandemic influenzaviruses [17]. The HA protein is the main antigenic stimulator of theacquired immunity [18] and is the major target for virus-neutralizingantibodies [19]. The prominent immune response against influenzaHA is presumed to be conducted to the head of the glycoprotein(amino acids 52-277) particularly against determined antigenic zonesthat enclose the receptor binding site [20]. It is well documented thatantibodies against HA in the circulation protect against intensiveviral pneumonia following the secretion of antibodies from the bloodinto the lungs of mice [21,22].
Because of the production of vaccines and new anti-viraldrugs are time-consuming, passive immunization by neutralizingantibodies provides urgent treatment for influenza pandemicemergency, particularly, for acutely exposed individuals [23].Neutralizing monoclonal antibodies successfully applied for therapyand prophylaxis of viral infection [7,24]. Due to HAMA (human antimouseimmunoglobulin antibody) responses induced by monoclonalantibodies, limited protections are reported [25,26]. Single chainfragment variable are small fragments (~25-30 kDa) comprised ofvariable heavy (VH) and light chain (VL) of antibody, which arejoined with a flexible peptide linker contains 15-20 amino acids [27].Human scFv antibodies have advantageous properties including:human origin, low molecular weight, higher tissue penetration,lower retention times in non desired tissues, better clearanceof theimmunocomplexes, preservation the binding specificity of the intactmonoclonal antibody and minimally immunogenic. Large quantitiesand cost-effective production of scFv antibodies in bacteria is possible[28-30]. Many scFvs are used successfully in clinical trial and treatment[30,31]. Highly specific scFv antibodies already produced to severalimportant pathogens, for example, against H5N1 Influenza viruses[32,33], Urease and lysate of H.pylori bacteria [34], p24 of HIV-1[35], thermolabile hemolysin (TLH) of Vibrio parahemolyticus [36],Varicella-zoster virus [37] and Human Papillomavirus type 16 [38].
In the present study, a phage antibody display library of singlechain variable fragments (scFv) was utilized for isolation scFvantibodies against the conserved sequence of the hemagglutinin of2009 pandemic H1N1 influenza virus. The antigenic specificities ofthese antibodies were evaluated by enzyme-linked immunosorbentassay (phage ELISA).
Materials and Methods
scFvs selection
A phage antibody display library of scFv was developed as described previously [39]. Phage clones displaying scFv were selectedfrom the library after four rounds of panning. Briefly, immunotubes(Nunc, Roskilde, Denmark) were coated with peptides as epitopes(amino acids 173-181 and 227-239 of hemagglutinin of influenzavirus at 4°C overnight. The phage-rescued supernatant (1010 PFU/mL) diluted with blocking solution was added to the tubes andincubated for 1h at room temperature. The recombinant phagemidwas introduced into competent E. coli TG1 cells, and rescued withhelper phage M13KO7 (Amersham, Biosciences). Four rounds ofpanning were performed. To confirm the existence of VH-Linker-VLinserts, PCR was performed on the clones obtained after panning.DNA fingerprinting of the scfv from individual E. coli clones weredetermined by using MvaI restriction enzyme (Roche DiagnosticGmbH, Mannheim, Germany). The common patterns were revealedby electrophoresis. One clone with the most frequent patternwas selected against each epitope and phage-rescued for furtherevaluations.
Determination of phage-antibody concentration
For determination of phage-antibody concentration of eachrescued phage, phage antibody supernatant was added to 1ml oflog phase TG1 E. coli and incubated with shaking at 37°C for 1 h.Serial dilution of bacteria was prepared and cultured on to 2TY Agar/Ampicillin medium at 30°C overnight. Number of clones per dilutionwas determined and phage concentration titer per milliliter wascalculated.
Evaluation of reactivity of scFvs by phage ELISA
Specificity of individual scFv was assessed by phage ELISA. The96 well ELISA plate was coated with peptides (dilution: 100 μg/ml inPBS) at 4°C overnight. An unrelated peptide was used as a negativecontrol. The wells were blocked with 3% skimmed milk for 2 h at 37°C.Following washing with PBS/Tween 20 and PBS, the phage-rescuedsupernatant of each clone containing the selected scFvs was added tothe wells. M13KO7 helper phage was added to peptide coated wellsas a negative antibody control. After incubation and washing, antifdbacteriophage antibody (Sigma, UK) was added and incubated for1hr. Following washing, HRP-conjocated anti-Rabbit IgG (Sigma,UK) antibody was added and left at room temperature for 1 h. 100μlof TMB solution was added to each well and the reaction was stoppedwith 100 μl of 1 M Sulfuric acid. The optical density of each well wasdetected at 450 nm by ELISA reader (BP-800, Biohit, USA).
Statistical analysis
Mann- Whitney test was used to compare the mean ratio of thephage ELISA results of phage display scFvs and of the control (nopeptide).
Results
Anti- hemagglutinin of 2009 pandemic H1N1 influenza Avirus scFvs
Figure 1 shows DNA-Fingerprinting of 20 panned clones againstpeptide 1(A) and peptide 2(B). Two common patterns obtainedagainst each peptide, scFv-I and svFv-II with the frequencies of 25%(5/20) and 55% (11/20) against peptide 1 and scFv -I’ and scFv II’with the frequencies of 20% (4/20) and 75% (15/20) against peptide 2. The common patterns are shown by common symbol in Figure 1.Lanes (1, 5, 8, 9, 17) and lanes (2, 3, 6, 7, 10, 11, 12, 13, 16, 19, 20)show the common patterns of scFv-I and scFv-II against peptide 1,respectively. Lanes (3, 8, 9, 17) and lanes (1, 2, 4, 5, 6, 7, 10, 12, 13, 14,15, 16, 18, 19, 20) represent common patterns of scFv-I’ and scFv-II’ which were selected against peptide 2, respectively.
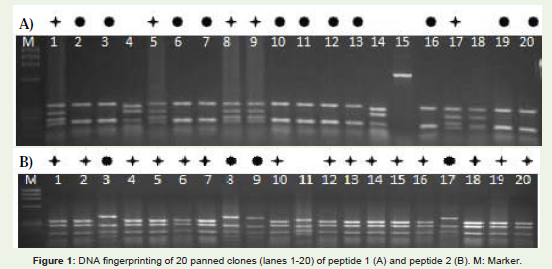
Figure 1: DNA fingerprinting of 20 panned clones (lanes 1-20) of peptide 1 (A) and peptide 2 (B). M: Marker.
Phage ELISA
The binding specificity of the selected scFvs to the related peptideswas measured by phage ELISA assay. All of four isolated scFvs (scFvI, scFvII, scFvI’, scFvII’) were found to bind with their related peptides(Figure 2). The absorbance obtained for reaction of scFv antibodieswith related peptide were significantly higher than the wells withno peptide (P value< 0.05). The M13KO7 helper phage showed noreactivity to the peptides.
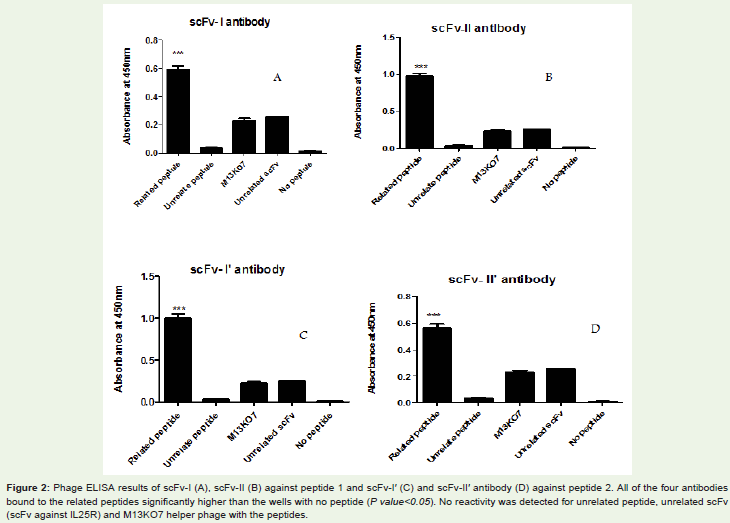
Figure 2: Phage ELISA results of scFv-I (A), scFv-II (B) against peptide 1 and scFv-I′ (C) and scFv-II′ antibody (D) against peptide 2. All of the four antibodiesbound to the related peptides significantly higher than the wells with no peptide (P value< 0.05). No reactivity was detected for unrelated peptide, unrelated scFv(scFv against IL25R) and M13KO7 helper phage with the peptides.
Discussion
One of the major causes of the global mortality is Influenzainfection and the infection remains a consistent health problem. Noimpressive prevention against Influenza infection has been identifiedtill now. Immunotherapy with antibodies is a helpful tool in thetreatment of infectious disease. Immunothreapy with neutralizingantibodies has been utilized against various viral infections includingprotection against Respiratory syncytial virus in cotton rats [40],prophylactic effect in experimentally infected chimpanzees againstHepatitis C virus [41], protection in mice and Guinea pigs inoculatedwith the rabies virus [42] and immunity against influenza viruses[43,44]. A strategy for defense against various viral infections is theproduction of antibodies to protective epitopes. Detection of theseepitopes is the main step in the development of effective antibodies.Between the 11 proteins that are encoded by gene segments ofinfluenza A virus, hemagglutinin is the major target of neutralizingantibodies as it is responsible for the attachment of the virus to thehost cells and determines the virulence of influenza virus. Conservedregions of hemagglutinin of influenza virus are principal targets forimmunotherapy against the virus. Highly conserved amino acids 173-181 and 227-239 were recognized in hemagglutinin of 2009 pandemicH1N1 influenza A virus [15,16,45]. These epitopes have beenidentified via B-1 and D-1 human monoclonal antibodies, respectively[16]. Considering the advantages of the scFv antibodies over intactantibodies, in the present study this format of antibody was used fortargeting of these epitopes. We chose peptide EGRMNYYWTLVEP(amino acids resides 227-239) of hemagglutinin fragmentsGKEVLVLWG (amino acides residues 173-181) and H1N1 InfluenzaA virus as immunodominant epitopes. Hemagglutinin fragmentsGKEVLVLWG (amino acids residues 173-181) and H1N1 Influenza A virus as immunodominant epitopes. Maneewatch et al. [33]isolated HuscFv antibodies against hemagglutinin of H5N1 influenzavirus. They reported that two of the selected HuscFvs had the highestneutralization effect against different strain of the H5N1 virus inMDCK culture and one HuscFv could rescue C57BL/6 mice thatwere infected with H5N1 influenza virus intranasally. In the previousstudies, the selection of scFv antibodies against the related peptidewas done by panning procedure. A number of specific scFvs have beenselected against key antigens using panning process [36,46,47]. In thisstudy we isolated four scFv antibodies (scFv-I, scFv-II, scFv-I’, scFv-II’) against peptide 1 and peptide 2 of hemagglutinin of influenza virus.The binding specificity of scFv antibodies to their related peptideswere measured by phage ELISA Assay. Results demonstrated thatthe selected scFvs against peptide 1 and 2 specifically bound to thecorresponding peptides compared to controls (Figure 2). The meanabsorbance of scFv-I, scFv-II and scFv-I’, scFv-II’ in reaction with thecorresponding peptides were significantly higher than the wells withno peptide, unrelated scFv, unrelated peptide and M13KO7 (P.value< 0.05). This represented the specificities of the four selected scFvs for the related peptides.
Immunotherapy via neutralizing antibodies against all speciesof influenza virus might be an ideal alternative for individuals whoare at high risk. This group includes: newborns, the old people andthe Immunodeficient patients who active vaccination probably doesnot help them. Neutralizing scFvs against hemagglutinin of influenzavirus would yield a new tool for therapy of influenza infection sinceHA has a principal role in the advent of human pandemic influenzaviruses. The well-known properties of scFv antibodies, speciallyhuman origin and wide penetration to the targets have made theseantibodies more applicable in immunotherapy. The high affinityspecific scFv antibodies against highly conserved neutralizingsequences of hemagglutinin of influenza were isolated in thisstudy suggest further evaluation of these antibodies to select highneutralizing scFvs to prevent against influenza infection in high riskindividuals.
Acknowledgment
The authors would like to acknowledge Shiraz University ofMedical Sciences for financial support. The present paper wasextracted from the Msc thesis written by Samaneh Alizadeh grant No:6713.
References
- Fox Jp, Cooney Mk, Hall Ce, Foy Hm (1982) Influenzavirus infections in Seattle families, 1975-1979. II. Pattern of infection in invaded households and relation of age and prior antibody to occurrence of infection and related illness. A J Epidemiology 116: 228-242.
- Johnson NP, Mueller J (2002) Updating the accounts: global mortality of the 1918-1920" Spanish" influenza pandemic. Bull Hist Med 76: 105-115.
- Berhane Y, Kehler H, Handel K, Hisanaga T, Xu W et al. (2012) Molecular and antigenic characterization of reassortant H3N2 viruses from Turkeys with a unique constellation of pandemic H1N1 internal genes. PLoS One 7: e32858.
- Cox NJ, Tamblyn SE, Tam T (2003) Influenza pandemic planning. Vaccine 21: 1801-1803.
- Peiris JS, Poon LL, Guan Y (2009) Emergence of a novel swine-origin influenza A virus (S-OIV) H1N1 virus in humans. Journal of Clinical Virology 45: 169-173.
- Shinde V, Bridges CB, Uyeki TM, Shu B, Balish A, et al. (2009) Triple-reassortant swine influenza A (H1) in humans in the United States, 2005–2009. N Engl J Med 360: 2616-2625.
- Shembekar N, Mallajosyula VVA, Mishra A, Yeolekar L, Dhere R, et al. (2013) Isolation of a High Affinity Neutralizing Monoclonal Antibody against 2009 Pandemic H1N1 Virus That Binds at the ‘Sa’Antigenic Site. PLoS One 8: e55516.
- Pebody R, McLean E, Zhao H, Cleary P, Bracebridge S, et al. (2010) Pandemic Influenza A (H1N1) 2009 and mortality in the United Kingdom: risk factors for death, April 2009 to March 2010. Euro Surveill 15: 19571.
- Jamieson DJ, Honein MA, Rasmussen SA, Williams JL, Swerdlow DL, et al. (2009) H1N1 2009 influenza virus infection during pregnancy in the USA. The Lancet 374: 451-458.
- Carlson A, Thung SF, Norwitz ER (2009) H1N1 influenza in pregnancy: what all obstetric care providers ought to know. Reviews in obstetrics and gynecology 2: 139-145.
- Tanaka T, Nakajima K, Murashima A, Garcia-Bournissen F, Koren G, et al. (2009) Safety of neuraminidase inhibitors against novel influenza A (H1N1) in pregnant and breastfeeding women. CMAJ 181: 55-58.
- Skehel JJ, Wiley DC (2000) Receptor binding and membrane fusion in virus entry: the influenza hemagglutinin. Annu Rev Biochemistry 69: 531-569.
- Knossow M, Skehel JJ (2006) Variation and infectivity neutralization in influenza. Immunology 119: 1-7.
- Hu W (2010) Highly conserved domains in hemagglutinin of influenza viruses characterizing dual receptor binding. Natural Science 2.
- Kubota-Koketsu R, Mizuta H, Oshita M, Ideno S, Yunoki M, et al. (2009) Broad neutralizing human monoclonal antibodies against influenza virus from vaccinated healthy donors. Biochem Biophys Res Commun 387: 180-185.
- Yamashita A, Kawashita N, Kubota-Koketsu R, Inoue Y, Watanabe Y, Ibrahim MS, et al (2010) Highly conserved sequences for human neutralization epitope on hemagglutinin of influenza A viruses H3N2, H1N1 and H5N1: Implication for human monoclonal antibody recognition. Biochem Biophys Res Commu 393: 614-618.
- DuBois RM, Aguilar-Yañez JM, Mendoza-Ochoa GI, Oropeza-Almazán Y, Schultz-Cherry S, et al. (2011) The receptor-binding domain of influenza virus hemagglutinin produced in Escherichia coli folds into its native, immunogenic structure. J virol 85: 865-872.
- Knipe D, Howley P, Griffin D, Lamb R, Martin M, Roizman B (2001) Fields virology, vol. 1. Philadelphia (EUA): Lippincott Williams & Wilkins.
- Martinez O, Tsibane T, Basler CF (2009) Neutralizing anti-influenza virus monoclonal antibodies: therapeutics and tools for discovery. Int Rev Immunology 28: 69-92.
- Ekiert DC, Friesen RH, Bhabha G, Kwaks T, Jongeneelen M, et al. (2011) A highly conserved neutralizing epitope on group 2 influenza A viruses. Science 333: 843-850.
- Ekiert DC, Bhabha G, Elsliger M-A, Friesen RH , Jongeneelen M, et al. (2009) Antibody recognition of a highly conserved influenza virus epitope. Science 324: 246-251.
- Sui J, Hwang WC, Perez S, Wei G, Aird D, et al. (2009) Structural and functional bases for broad-spectrum neutralization of avian and human influenza A viruses. Nat Struct Mol Biol16: 265-273.
- Keller MA, Stiehm ER (2000) Passive immunity in prevention and treatment of infectious diseases. Clin Microbiol Rev 13: 602-614.
- Hessell A, Rakasz EG, Tehrani DM, Huber M, Weisgrau KL, Landucci G, et al (2010) Broadly neutralizing monoclonal antibodies 2F5 and 4E10 directed against the human immunodeficiency virus type 1 gp41 membrane-proximal external region protect against mucosal challenge by simian-human immunodeficiency virus SHIVBa-L. J virol 84: 1302-1313.
- Gruber R, van Haarlem LJ, Warnaar SO, Holz E, Riethmüller G (2000) The human antimouse immunoglobulin response and the anti-idiotypic network have no influence on clinical outcome in patients with minimal residual colorectal cancer treated with monoclonal antibody CO17-1A. Cancer Res 60: 1921-1926.
- Pucca MB, Bertolini TB, Barbosa JE, Galina SVR, Porto GS (2011) Therapeutic monoclonal antibodies: scFv patents as a marker of a new class of potential biopharmaceuticals. Brazilian Journal of Pharmaceutical Sciences 47: 31-38.
- Ahmad ZA, Yeap SK, Ali AM, Ho WY, Alitheen NBM, et al. (2012) scFv antibody: principles and clinical application. Clin Dev Immunol 2012.
- Deckert P (2009) Current constructs and targets in clinical development for antibody-based cancer therapy. Curr Drug Targets 10: 158-175.
- Kobayashi N, Odaka K, Uehara T, Imanaka-Yoshida K, Kato Y, et al. (2011) Toward in vivo imaging of heart disease using a radiolabeled single-chain Fv fragment targeting tenascin-C. Anal Chem 83: 9123-9130.
- Holliger P, Hudson PJ (2005) Engineered antibody fragments and the rise of single domains. Nat Biotechnol 23: 1126-1136.
- Cardinale A, Filesi I, Vetrugno V, Pocchiari M, Sy M-S, et al. (2005) Trapping prion protein in the endoplasmic reticulum impairs PrPC maturation and prevents PrPSc accumulation. J Biol Chem 280: 685-694.
- Yang J, Yoshida R, Kariya Y, Zhang X, Hashiguchi S, et al. (2010) Characterization of human single-chain antibodies against highly pathogenic avian influenza H5N1 viruses: mimotope and neutralizing activity. J Biochem 148: 507-515.
- Maneewatch S, Thanongsaksrikul J, Songserm T, Thueng-In K, Kulkeaw K, et al. (2009) Human single-chain antibodies that neutralize homologous and heterologous strains and clades of influenza A virus subtype H5N1. Antivir Ther 14: 221-230.
- Reiche N, Jung A, Brabletz T, Vater T, Kirchner T, Faller G (2002) Generation and characterization of human monoclonal scFv antibodies against Helicobacter pylori antigens. Infect Immun 70: 4158-4164.
- Obregon P, Chargelegue D, Drake PM, Prada A, Nuttall J, et al. (2006) HIV‐1 p24-immunoglobulin fusion molecule: a new strategy for plant‐based protein production. Plant Biotechnol J 4: 195-207.
- Wang R, Fang S, Wu D, Lian J, Fan J, et al. (2012) Screening for a Single-Chain Variable-Fragment Antibody That Can Effectively Neutralize the Cytotoxicity of the Vibrio parahaemolyticus Thermolabile Hemolysin. Appl Environ Microbiol 78: 4967-4975.
- Kausmally L, Waalen K, Løbersli I, Hvattum E, Berntsen G, Michaelsen T, et al (2004) Neutralizing human antibodies to varicella-zoster virus (VZV) derived from a VZV patient recombinant antibody library. J Gen virol 85: 3493-3500.
- Donà MG, Giorgi C, Accardi L (2007) Characterization of antibodies in single-chain format against the E7 oncoprotein of the human papillomavirus type 16 and their improvement by mutagenesis. BMC cancer 7: 25.
- Nejatollahi F, Malek-Hosseini Z, Mehrabani D (2008) Development of single chain antibodies to P185 tumor antigen. Iranian Red Crescent Medical Journal 10: 298-302.
- Prince GA, Hemming VG, Horswood RL, Baron PA, Chanock RM (1987) Effectiveness of topically administered neutralizing antibodies in experimental immunotherapy of respiratory syncytial virus infection in cotton rats. J virol 61: 1851-1854.
- Lorenzo CD, Angus AG, Patel AH (2011) Hepatitis C virus evasion mechanisms from neutralizing antibodies. Viruses 3: 2280-2300.
- Both L, Banyard AC, van Dolleweerd C, Horton DL, Ma JK, et al. (2012) Passive immunity in the prevention of rabies. The Lancet infectious diseases12: 397-407.
- Yoshida R, Igarashi M, Ozaki H, Kishida N, Tomabechi D, et al. (2009) Cross-protective potential of a novel monoclonal antibody directed against antigenic site B of the hemagglutinin of influenza A viruses. PLoS pathog 5.
- Prabhu Niang J, Prabakaran M, Ho H-T, Velumani S, Qiang J, et al. (2009) Monoclonal antibodies against the fusion peptide of hemagglutinin protect mice from lethal influenza A virus H5N1 infection. J virol 83: 2553-2562.
- Moattari A, Ashrafi H, Kadivar M, Kheiri M, Shahidi M, et al. (2010) Antigenic variations of human influenza virus in Shiraz, Iran. Indian J Med Microbiol 28: 114.
- Nejatollahi F, Hodgetts SJ, Vallely PJ, Burnie JP (2002) Neutralising human recombinant antibodies to human cytomegalovirus glycoproteins gB and gH. FEMS Immunol Med Microbiol 34: 237-244.
- Nejatollahi F, Abdi S, Asgharpour M (2013) Antiproliferative and Apoptotic Effects of a Specific Antiprostate Stem Cell Single Chain Antibody on Human Prostate Cancer Cells. J oncol 2013.