Review Article
Relationship between Maternal Aging and Risk of Chromosome 21 Nondisjucntion: Where We Are and Where We Have To Go?
Papiya Ghosh2, Sujay Ghosh1*
Corresponding author: Dr. Sujay Ghosh, Cytogenetics & Genomics Research Unit, Department of Zoology, University of Calcutta, 35 Ballygunge Circular Road, Kolkata, West Bengal India, Pincode 00019, E-mail: g_sujoy@yahoo.com
Citation: Ghosh P, Ghosh S. Relationship between Maternal Aging and Risk of Chromosome 21 Nondisjucntion: Where We Are and Where We Have To Go? J Cell Sci Molecul Biol. 2015;2(1): 114.
Copyright © 2015 Sujay Ghosh et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Submission: 04/09/2015; Accepted: 21/09/2015; Published: 26/09/2015
Summary
Several lines of researches have revealed that the women having Down Syndrome (DS) child are usually biologically older than the women of samechronological age with euploid baby. On other hand, Women having DS child often suffer from early onset of Alzheimer disease (AD). Frequent associationbetween AD and DS in some families intuitively suggests sharing of some common genetic risk factors between maternal precocious aging and chromosomalnondisjunction. Subsequent molecular analyses have revealed polymorphisms of certain genes simultaneously associated with AD and DS birth. In thisregard polymorphic alleles of APOE, Presinilin-1 and BubR1 are of particular interest. So, understanding the molecular relation between accelerated maternalaging and increased risk of DS conception will provide basis of future research that may have large medical applicability.
Keywords: Down syndrome; Maternal age; Accelerated aging; Nondisjunction; Telomere
Introduction
The aetiology of nondisjunction (NDJ) of Chromosome 21(Ch21) that causes birth of baby with Down syndrome (DS) isan unsolved issue in the field of medical genetics for decades. DSrepresents the most frequent genetic form of intellectual disabilityin human. In overwhelming majority of live born DS cases NDJerrors originate in oocyte and the frequency of error increaseswith advancing chronological age of women [1,2]. The associationbetween advanced maternal age of conception and elevated riskof DS birth was reported initially by Penrose [3] and subsequentlyconfirmed in other studies [4-6]. On other hand, population basedstatistical analyses have revealed elevated risk of Alzheimer Disease(AD), (which is the expression of aging associated dementia) among some women who bear DS child before 35 years of age; butthe mothers who have DS pregnancy beyond 35 years of their agedo not exhibit such association [7]. Thus the relationship betweenmaternal age and risk of Ch21 NDJ in oocyte is intriguing and can beviewed in two different but parallel ways; firstly, the gradual increasein the risk of NDJ with advancing maternal chronological age andsecondly, stochastic risk among chronologically younger women whootherwise suffer from accelerated biological aging. This second notionis truly riddling as our current understanding of maternal biologicalaging and its relation with the risk of Ch21 NDJ at molecular levelis surprisingly limited. The present review is focused to address theprospective genetic and epidemiological candidates that may relateaccelerated aging and increased risk among women to conceive DSchild.
Chronological Aging and Increasing Risk of NDJ
The hypotheses that explain association of advancing maternalage with NDJ have generalized approach to accommodate all typesof human aneuploidy involving autosomes and sex chromosome.Several hypotheses have been put forward to elucidate the link betweenadvancing maternal age and higher incidence of aneuploid oocyteformation, though no one has proved to be completely satisfactory.The most popular hypothesis [8] holds that protracted tenure ofoogenesis interrupted with meiotic halts probably makes the eggsmore vulnerable to the aging effect than sperms (Text Box 1). This longperiod of oocyte maturation causes deteriorative changes to accrueover time either in the oocyte or its milieu. Examples of such factorsare diminishing amount of a meiotic proteins, like those maintainingsister chromatid adhesion [9] or meiotic checkpoint components [10]or weakening of centromere cohesion due to age-related reduction incentromere associated proteins MCAK [11]. This list of age relatedrisks may also include the accumulation of environmentally induceddamages to the meiotic machinery over time or genetic changes suchas mitochondrial deletions [12]. Among all these variables the spindleassembly check point (SAC) components and sister chromatidcohesion (SCC) were investigated thoroughly [13]. The SAC is amolecular machine that ensures proper chromosome separation inboth mitosis and meiosis. In meiosis SAC prevents anaphase untilall chromosomes get attached properly to the spindle. The SACincludes MAD2L1, BUB1B, and TTK [14,15], which show decline inconcentration with age and causes misalignment of chromosomes onmetaphase plate in mouse [16]. This observation suggests that errorsin SAC function contribute to age-related aneuploidy. Consistentwith this hypothesis, disrupted spindles, misaligned chromosomesand decreased expression of SAC components MAD2L1 and BUB1Bhave evident in aged human oocytes [17,18]. On the other hand theSCC mediates physical pairing of duplicated chromosome whichis essential for their appropriate distribution in daughter cells. Thecohesion along chromosome arms keeps the bivalent intact in meiosisI (MI), and centromere cohesion holds sister chromatids togetherin MII. Defect in cohesion distal to crossover sites may result in ashift of chiasmata placement (chiasmata slippage) or even prematurebivalent separation in MI, whereas reduced centromere cohesion may result in premature separation of sister chromatids in MII [18]. Theloss of cohesion with maternal age for distally placed chisama [19]is consistent with the idea that cohesion defects may contribute toage related aneuploidy [20]. More specifically the meiotic cohesinproteins Smc1β and Rec8 exhibit age related reduction in mouseoocyte resulting in misaligned chromosomes and aneuploidy [20,21].
Another component that is supposed to decline with age andcontributes significantly to the maternal aging effect on DS birth is thesurveillance system of ovary that ensures segregation of achiasmatechromosomes at meiotic anaphase [1]. Chiasma formation andsubsequent recombination are prerequisite of faithful separation ofhomologous chromosomes at meiotic anaphase (Text Box 2). Absenceof chiasma, faulty chiasma configurations and reduction in chiasmafrequency result in NDJ of Ch21 and subsequent DS birth [22]. Ahigh proportion of achiasmate Ch21 tetrad was reported among themothers of DS having age >35 year [1]. As the decision regardingchiasma formation is taken in foetal ovary, high frequency of achisamate nondisjoined Ch21 in older oocyte can only be explainedby down regulation of surveillance system. Human proteins involvedin proper segregation of nonexchange chromosome show downregulation with increasing ovarian age [18,23].
Advanced Biological Aging among DS bearing Women
All the above mentioned hypotheses that incriminate decay insubcellular protein components in oocyte for age-related increase oftrisomy 21(T21) conception do consider the maternal chronologicalage, although individual variation in the extent of these changescould occur [5]. A new idea regarding maternal age effect on meioticerrors was proposed by Warburton, [24,25] and the hypothesisstates that the “biological aging” or “ovarian aging” is the cause ofincreasing rate of meiotic errors. The central theme of this hypothesisis the prediction that biological aging is different among womenof the same chronological age, and that the frequency of trisomicconceptions depends upon the biological age of the woman ratherthan their chronological age [25]. The biological age of women canusually be assessed by counting the falling number of antral follicleswith chronological age together with decrease in total oocyte poolsize [26,27]. These altogether alter the optimum hormonal balance inovary, which is marked by falling concentration of serum inhibin Aand B, decline in estrogens surge and elevated level of FSH [25]. Thischange in hormone balance is related to increased rate of aneuploidyat advanced maternal age. Support to this prediction is available fromthe experiment in mouse model [28].
The “biological aging” hypothesis predicts that women with atrisomic conception should on the average have an older “ovarianage” than women of the same chronological age with normaleuploid conception [25] and women having T21 pregnancy haveaverage earlier (~1 year) age of menopause [29] than the womenwith chromosomally normal conception. If this were the case, onewould expect that after a trisomic conception, the risk of a subsequenttrisomy birth for any chromosome should be higher than the maternalage-related risk. Support to this prediction has been provided fromthe observation that risk of trisomic pregnancy after a previoustrisomic conception is about 1.7 times the maternal age-related risk[30]. Mathematic model proposed by Kline and Levin [31] estimatedthat women with trisomy pregnancy experience 0.9 years earlymenopause which suggests women with trisomy pregnancy sufferfrom advanced ovarian age than the women with chromosomallynormal pregnancies. Population sample survey for estimating medianage of menopause among the women with trisomic pregnancy alsosuggested an early cessation of menstrual cycle among them thanthe mothers with chromosomally normal foetus [29]. Elevated levelof FSH has been reported among the women with DS pregnancy[32,33], which suggests precocious aging among them.
Owing to lack of conclusive evidence in favour of the ‘biologicalaging, Warburton [25] proposed an alternative notion, namely‘limited oocyte pool hypothesis’ which stated that with increasingbiological age gradual decrease in the number of antral follicleleaves only the premature or post mature oocyte to ovulate andthat inevitably leads to aneuploid conception. Recently Kline et al.[34] conducted a survey on hormonal level of women with trisomicpregnancy and the outcome of the study supported the “reduced oocyte pool hypothesis”. The authors inferred that some womenhave smaller follicle content than the others of same chronologicalage and the former group are susceptible for rapid ovarian aging andassociated trisomic conceptions. All these findings imply an intuitiveexistence of predisposing factors that make some women susceptiblefor both the accelerated aging and to have trisomic conception.
Accelerated Genetic Aging among DS bearing Women
If the biological aging hypothesis holds true, the question thatobviously comes in mind whether, any genetic factor underlies thecause of biological aging or not. In others words, some women maybe genetically predisposed to rapid molecular aging and thereforeexperience advanced biological and ovarian aging, which increasesthe chance of trisomic conception. Attempt to resolve this issue wasstarted by measuring the telomere length (TL) of women havingtrisomic pregnancy as the telomeric repeat attrition is an authenticmarker of molecular aging in somatic tissue [35,36]. The initial studyfor determining the maternal molecular age for DS was carriedby Dorland et al. [37] who estimated the TL of peripheral bloodlymphocytes from women having DS baby and compared the TL withthat of women having euploid child. This study included women of< 35 years age and did not achieve statistically significant differencebetween cases and controls. The outcome of this study suggestedthat the younger women who have DS baby do not differ in theirgenetic or molecular age from the women of similar chronologicalage having euploid baby. A positive correlation between loss of TLin somatic tissue and shorter reproductive life span due to ovarianaging was reported in other studies [38,39]. The study on the womenwith history of recurrent miscarriages of trisomic foetus conductedby Hanna et al. [40] revealed no difference in TL of lymphocytestaken from cases and controls. In contrast to this report, a very recentstudy on DS bearing mothers [41] have demonstrated a significantdifference in TL among the case and control women of older age (>34years). In this cross sectional study the participating women werestratified initially by age of conception (young :< 29 years, middle:29-34 and old >34 years) and then by meiotic origin of NDJ of Ch21(MI NDJ or MII NDJ for DS case category). The TL of lymphocyteswas estimated by restriction digestion-Southern blot approach. Theresult shows much faster rate of TL shortening among mothers ofDS child and the rate of shortening is significantly different from theestimated TL attrition rate of controls (61 bp/years in control; 111bp/year in MI NDJ group; 230 bp/year in MII NDJ group) (Figure 1).Moreover, authors found significant difference in mean TL amongthe controls, MI and MII (p = 0.004 for MI-NDJ vs. MII-NDJ, p < 0.001 for MI-NDJ vs. control and p < 0.001 for MII-NDJ vs. control) with shortest TL recorded in older MII group (Figure 2). The authorsproposed ‘genetic aging hypothesis’ which holds the notion thatsome women are predisposed to rapid genetic and molecular agingand its effect is exacerbated at advance age when age-related naturaldeteriorative changes also affect the chromosome separation systemleading to NDJ. The notion suggested some intuitive link betweentelomere maintenance system (i.e., system of molecular aging) andchromosome segregating apparatus at molecular level possibly insharing one or more molecular components. Some polymorphisms oftelomere maintenance components TERC and TERT are reported [42-44] to associate the rapid telomere attrition and some cardiovascular and cerebro-vascular phenotypes resembling aging [45], but their rolehave not been evaluated as risk factors for Ch21 NDJ and DS birth.
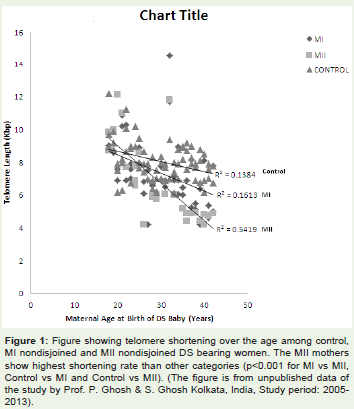
Figure 1: Figure showing telomere shortening over the age among control,MI nondisjoined and MII nondisjoined DS bearing women. The MII mothersshow highest shortening rate than other categories (p< 0.001 for MI vs MII,Control vs MI and Control vs MII). (The figure is from unpublished data ofthe study by Prof. P. Ghosh & S. Ghosh Kolkata, India, Study period: 2005-2013).
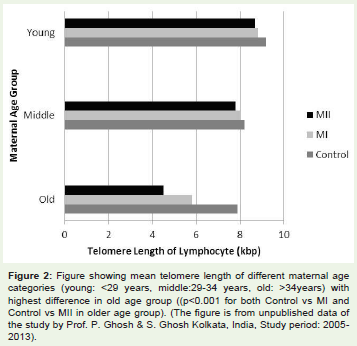
Figure 2: Figure showing mean telomere length of different maternal agecategories (young: < 29 years, middle:29-34 years, old: >34years) withhighest difference in old age group ((p< 0.001 for both Control vs MI andControl vs MII in older age group). (The figure is from unpublished data ofthe study by Prof. P. Ghosh & S. Ghosh Kolkata, India, Study period: 2005-2013).
Signature for Accelerated Aging: Early onset of ADamong DS bearing women
Probably the most convincing evidence for accelerated agingamong women having DS child is the early onset Alzheimer disease (AD) among them. Several studies have suggested the possibilityof shared risk of AD and DS among women. In their study on DSfamilies, Heston et al. [46] noted an association of early age at onsetof AD among the family members and inheritance was throughmaternal line. The study by Schupf et al. [7] revealed that womenhaving DS baby at < 35 years age suffered from early onset of AD andthey hypothesized that DS and AD shared a genetic susceptibilitythrough accelerated aging of mothers. The authors estimated therelative risk of developing symptoms of dementia among women ofage < 35 years bearing DS child and it were about five times than forcontrols. In search of putative candidates that link the early onset ofAD phenotypes among women and their DS conception, differentgroups of workers have reported the allelic variants of two genes. Thefirst of them is ε4 allele of APOE gene (located on Chromosome19)and the other one is intronic polymorphism of the gene Presenilin-1(located on Chromosome14). The study by Hofman et al. [47]suggested an association between meiotic stage of NDJ and theAPOE ε4 allele among mothers who gave birth to a DS child when32 years of age and the authors inferred that the shared susceptibilityto DS and AD might be mediated by an increased frequency of the ε4allele. The study revealed that frequency of the ε4 allele was greaterin young mothers with MII-NDJ than in young mothers with a MINDJ(30% versus 19%), but did not differ between young motherswith meiosis I error and control mothers (19% versus 17%). Thisobservation was in agreement with the findings of Avramopoulos etal. [48] who hypothesized that APOE allele ε4 could be a risk factorfor chromosomal NDJ and probably links the meiotic error with earlyonset of aging phenotypes among chronologically younger women.Beside APOE/ ε4 analyses, Petersen et al. [49] reported an associationamong MII-NDJ, Down syndrome conception and preferentialoccurrence of intron 8 polymorphism of Presenilin 1 gene. ThePresenilin-1 is a candidate for kinetochore functioning complex,involved in chromosome segregation and mutant form of this geneis associated with early onset of autosomal dominant AD phenotype[49]. The study of Petersen et al. [49] included 168 families eachwith free trisomy 21 child and the authors found a higher frequencyof susceptible Presinilin-1 intronic polymorphism (allele 1) in homozygous state among the mothers experienced MII error (nearly53%, P< 0.05). More interestingly, the authors reported significantco-segregation of Presenilin-1 intronic allele 1 and APOE allele ε4allele among the DS child bearing mothers (68%) when comparedwith non- ε4 allele bearing mothers (P< 0.01). The AD susceptiblepolymorphic alleles of Presenilin-1 and APOE exhibited increasedfrequency of occurrence among young mothers (< 32 years age of DSconception) with MII errors [48,49]. To refute this finding Bhowmiket al. [50] have conducted analyses on Indian women and confirmedthat Presenilin-1 polymorphism is associated with MII NDJ amongthe young mother. This study included 96 probable Alzheimer’spatients (mean age 62.26 ± 11.93 years with 70.48% male and 29.52%female subjects), 173 age matched healthy controls and 136 DSfamilies. The authors reported a rare polymorphism rs201992645[(NG_007386.2:g.66696T>A or NM_000021.3:c.868+37T>A)ss515119316] within the intron 8 of PSEN-1 gene at the 73664874thnucleotide position on chromosome 14 (GRCh37.p5 Assembly). Thechange involves nucleotide transition T>A. The mutant ‘A’ alleleexists only as heterozygous condition (T/A) with genotype and allele frequencies of 0.031 and 0.02 respectively for AD cases and 0.029and 0.01 respectively for DS. Within the parents of DS, the mutantallele was recorded only among the mother with genotypic andallelic frequencies of 0.029 and 0.01 respectively. To anticipate theprobable damaging effect of the detected intronic mutation on PSEN-1 expression and functions, the authors employed bioinformaticssoftware which identified said polymorphic site caused alternation insplice site at intron exon junction that in turn may result into changein protein structure and functions. Result of this study inferredPresenilin-1 as prospective molecular link between AD and DS birthand it predisposes the women simultaneously for Ch21 NDJ and ADirrespective of maternal age of conception.
To explain the familial segregation of AD associated geriatric phenotypes and trisomy 21 birth, an alternative hypothesis has been put forward and the central idea of this notion is Ch21 mosaicismin the germ cells and brain accounts for familial segregation of ADand trisomy 21, [51]. Recently proposed ‘oocyte mosaicism selectionmodel’ [52] provides the evidence of presence of Ch21 disomicmosaic cells in the germinal epithelium of ovary and suggest is maybe the underlying cause of DS birth. But presence of chromosomallymosaic germinal cell line may not an inevitable evidence of existenceof such aneuploid cells in neuronal tissue. Therefore, the commongenetic origin risk of trisomy 21 conception and early onset of ADphenotype remain inexplicable.
Conclusion and Future Research
Understanding the cause of frequent co-occurrence of acceleratedgenetic aging and risk to conceive TS21 foetus among some women isdifficult. Several apparent conflicting evidences have made the picturemessy. It is now known that the cause of DS birth is multifactorial,includes both maternal age-dependent and age-independent risk[53] and there are several tiers of interactions among genetic factors,environmental agents and maternal age. But the efforts for findingout the origin of accelerated molecular aging and its connection withchromosomal NDJ is tantalizingly low. Sincere research works haveidentified some potential candidate genes that may be the ‘missinglink’ between the molecular aging mechanism and chromosomeseparation system and act as maternal age independent riskfactor for DS birth. The BubR1 is of particular interest as availableevidence [23] suggests its role for aging and associated chromosomalmissegregation. Moreover, the co-segregation of APOE allele ε4 andPresinilin-1 intronic polymorphism allele-1 and their role in originof trisomy21 may be the subject of future research. The discoveryof rapid shortening of TL among DS bearing mothers opens anotherline of thinking. Further researches are needed to explore the geneticpolymorphism of the molecular components of telomere thatmight predispose TL attrition on one hand and NDJ on the other.There are several cross-talks among different genetic circuits and asharing of genetic components between molecular aging process andchromosome segregation apparatus may not be overruled. Althoughthe possibilities are speculative, nevertheless, the available evidencesare suggestive of common shared genetics of accelerated biologicaland molecular aging of women and their risk to conceive trisomy 21foetus. Future researches are needed to realize explicitly what is goingexactly at subcellular level in the oocyte.
Financial Support: The work is supported by University GrantCommission (UGC), New Delhi; Sanction No.F.PS068/10-11.
Conflict of interest Statement: None declared.
Text Box 1 None declared.
Meiosis is initiated in the human fetal ovary at 11–12 weeks ofgestation, but becomes arrested after completion of homologouschromosome pairing and recombination. This meiotic-halt lastsfor several years until the elevated level of LH and FSH resume theprocess at onset of puberty. Then the oocyte completes meiosisI and enters meiosis II and again undergoes a phase of pause. Itcompletes the meiosis II after the sperm enter its cytoplasm followingfertilization. Thus, the oocyte, whose ovulation marks the menarchy,remains in pause for shortest period and that ovulate just precedingmenopause experiences longest period of arrest. This long tenure ofoocyte development makes it vulnerable to acquire environmentalhazards within its microenvironment which inevitably increases therisk of chromosomal NDJ at advanced age of women. Moreover,each menstrual cycle recruits several follicles to initiates growth.Subsequently, all follicles degenerate except one which becomes theGraafian follicle. The follicular atresia reduces the follicular pool inovary and left more defective oocyte to ovulate stochastically in moreadvancing age of women. This explains why increased rate of Downsyndrome birth is associated with advancing maternal age.
Text Box 2
Recombination between homologous chromosomes is anessential criterion for their proper segregation towards opposite pole.A chiasma at the middle of chromosome arms maintains and balancesthe pull of spindle fibre from the opposite poles which ensures properchromosome movement at anaphase. So, any deviation in chiasmaplacement on chromosome arm inevitably leads to malsegregation ofchromosomes. For chromosome 21 three defective exchange patternshave been observed. These include chiasma too close to centromere,too close to telomere and no chiasma formation. The peri-centromericchiasma cause chromosomal entanglement and oocyte compromisesthis configuration with increasing maternal age and ovulates with thisdefect. So this configuration is maternal age dependent risk factor forDS birth. On the other hand, telomeric chiasma formation involvesvery little sister chromatid adhesion protein recruitment leadingto premature sister chromatid separation and this is maternal ageindependent risk for DS birth. The chromosome 21 that does notexperience any chiasma formation segregates randomly and impartsrisk in maternal age independent manner.
References
- Oliver TR, Feingold E, Yu K, Cheung V, Tinker S, et al. (2008) New insights into human NDJ of chromosome 21 in oocytes. PLoS Genet 4: e1000033.
- Ghosh S, Feingold E, Dey SK (2009) Etiology of Down Syndrome: Evidence for Consistent Association among Altered Meiotic Recombination, NDJ and Maternal Age Across Populations. Am J Med Genet 149A: 1415-1420.
- Penrose LS (1954) Mongolian idiocy (mongolism) and maternal age. Ann N Y Acad Sci 57: 494-502.
- Lamb NE, Yu K, Shaffer J, Feingold E, Sherman SL (2005) Association between maternal age and meiotic recombination for trisomy 21. Am J Hum Genet 76: 91-99.
- Sherman SL, Allen EG, Bean LH, Freeman SB (2007) Epidemiology of Down syndrome. Ment Retard Dev Disabil Res Rev 13: 221-227.
- Ghosh S, Bhaumik P, Ghosh P, Dey SK (2010) Chromosome 21 non-disjunction and Down syndrome birth in an Indian cohort: analysis of incidence and aetiology from family linkage data. Genet Res (Camb) 92: 189-197.
- 7.Schupf N, Kapell D, Nightingale B, Mohlenhoff J, Bewley S, et al. (2001) Specificity of the fivefold increase in AD in mothers of adults with Down syndrome. Neurology 57: 979-984.
- Gondos B, Westergaard L, Byskov AG (1986) Initiation of oogenesis in the human fetal ovary: ultrastructural and squash preparation study. Am J Obstet Gynecol 155: 189-195.
- Hodges CA, Revenkova E, Jessberger R, Hassold TJ, Hunt PA (2005) SMC1beta-deficient female mice provide evidence that cohesins are a missing link in age-related nondisjunction. Nat Genet 37: 1351-1355.
- Garcia-Cruz R, Brieño MA, Roig I, Grossmann M, Velilla E, et al. (2010) Dynamics of cohesin proteins REC8,STAG3,SMC1 beta and SMC3 are consistent with a role in sister chromatid cohesion during meiosis in human oocytes. Hum Reprod 25: 2316-2327.
- Eichenlaub-Ritter U, Staubach N, Trapphoff T (2010) Chromosomal and cytoplasmic context determines predisposition to maternal age-related aneuploidy: brief overview and update on MCAK in mammalian oocytes. Biochem Soc Trans 38: 1681-1686.
- Van Blerkom J (2011) Mitochondrial function in the human oocyte and embryo and their role in developmental competence. Mitochondrion 11: 797-813.
- Chiang T, Schultz RM, Lampson MA (2011) Age-dependent susceptibility of chromosome cohesion to premature separase activation in mouse oocytes. Biol Reprod 85: 1279-1283.
- Hached K, Xie SZ, Buffin E, Cladière D, Rachez C, et al. (2011) Mps1 at kinetochores is essential for female mouse meiosis I. Development. 138: 2261-2271.
- Niault T, Hached K, Sotillo R, Sorger PK, Maro B, et al. (2007) Changing Mad2 levels affects chromosome segregation and spindle assembly checkpoint control in female mouse meiosis I. PLoS One 2: e1165.
- Pan H, Ma P, Zhu W, Schultz RM (2008) Age-associated increase in aneuploidy and changes in gene expression in mouse eggs. Dev Biol 316: 397-407.
- McGuinness BE, Anger M, Kouznetsova A, Gil-Bernabé AM, Helmhart W, et al. (2009) Regulation of APC/C activity in oocytes by a Bub1-dependent spindle assembly checkpoint. Curr Biol 19: 369-380.
- Steuerwald N, Cohen J, Herrera RJ, Sandalinas M, Brenner CA (2001) Association between spindle assembly checkpoint expression and maternal age in human oocytes. Mol Hum Reprod 7: 49-55.
- Subramanian VV, Bickel SE (2008) Aging predisposes oocytes to meiotic nondisjunction when the cohesin subunit SMC1 is reduced. PLoS Genet 4: e1000263.
- Chiang T, Schultz RM, Lampson MA (2012) Meiotic origins of maternal age-related aneuploidy. Biol Reprod 86: 1-7.
- Chiang T, Schultz RM, Lampson MA (2011) Age-dependent susceptibility of chromosome cohesion to premature separase activation in mouse oocytes. Biol Reprod 85:1279-1283.
- Lamb NE, Sherman SL, Hasold TJ (2005) Effect of meiotic recombination on production of aneuploid gametes in humans. Cytogenet Genome Res 111: 250-255.
- Baker DJ, Jeganathan KB, Cameron JD, Thompson M, Juneja S, et al. (2004) BubR1 insufficiency causes early onset of aging associated phenotypes and infertility in mice. Nat Genet 36: 744-749.
- Warburton D (1989) The effect of maternal age on the frequency of trisomy: change in meiosis or in utero selection? Prog Clin Biol Res 311: 165-181.
- Warburton D (2005) Biological aging and etiology of aneuploidy. Cytogenetics and Genome Res 111: 266-272.
- Scheffer GJ, Broekmans FJ, Dorland M, Habbema JD, Looman CW, et al. (1999) Antral follicle counts by transvaginal ultrasonography are related to age in women with proven fertility. Fertil Steril 2: 845-851.
- Kline J, Kinney A, Reuss ML, Kelly A, Levin B, et al. (2004) Trisomic pregnancy and the oocyte pool. Hum Reprod 19: 1633-1643.
- Roberts R, Iatropoulou A, Ciantar D, Stark J, Becker DL, et al. (2005) Follicle-stimulating hormone affects metaphase I chromosome alignment and aneuploidy in mouse oocytes matured in vitro. Biol Reprod 72: 107-118.
- Kline J, Kinney A, Levin B, Warburton D (2000) Trisomic pregnancy and earlier age at menopause. Am J Hum Genet 67: 395-404.
- Warburton D, Dallaire L, Thangavelu M, Ross L, Levin B, et al. (2004) Trisomy recurrence: a reconsideration based on North American data. Am J Hum Genet 75: 376-385.
- Kline J, Levin B (1992) Trisomy and age at menopause: predicted associations given a link with rate of oocyte atresia. Pediatr Perinat Epidemiol 6: 225-239.
- Nasseri A, Mukherjee T, Grifo JA, Noyes N, Krey L, et al.(1999) Elevated day 3 serum follicle stimulating hormone and/or estradiol may predict fetal aneuploidy. Fertil Steril 71: 715-718.
- van Montfrans JM, van Hooff MH, Martens F, Lambalk CB (2002) Basal FSH, estradiol and inhibin B concentrations in women with a previous Down’s syndrome affected pregnancy. Hum Reprod 17: 44-47.
- Kline JK, Kinney AM, Levin B, Kelly AC, Ferin M, et al. (2011) Trisomic pregnancy and elevated FSH: implications for the oocyte pool hypothesis. Hum Reprod 26: 1537-1550.
- Cawthon RM, Smith KR, O'Brien E, Sivatchenko A, Kerber RA (2003) Association between telomere length in blood and mortality in people aged 60 years or older. Lancet 361: 393-395.
- Njajou OT, Cawthon RM, Damcott CM, Wu SH, Ott S, et al. (2007) Telomere length is paternally inherited and is associated with parental lifespan. Proc Natl Acad Sci USA 104: 12135-121359.
- Dorland M, van Montfrans JM, van Kooij RJ, Lambalk CB, te Velde ER (1998) Normal telomere lengths in young mothers of children with Down’s syndrome. Lancet 352: 961-962.
- Aydos SE, Elhan AH, Tükün A (2005) Is telomere length one of the determinants of reproductive life span? Arch Gynecol Obstet 272: 113-116.
- Keefe DL, Liu L (2009) Telomeres and reproductive aging. Reprod Fertil 21: 10-14.
- Hanna CW, Bretherick KL, Gair JL, Fluker MR, Stephenson MD, et al. (2009) Telomere length and reproductive aging. Hum Reprod 24: 1206-1211.
- Ghosh S, Feingold E, Chakraborty S, Dey SK (2010) Telomere length is associated with types of chromosome 21 NDJ: a new insight into the maternal age effect on Down syndrome birth. Hum Genet 127: 403-409.
- Shen Q, Zhang Z, Yu L, Cao L, Zhou D, et al. (2011) Common variants near TERC are associated with leukocyte telomere length in the Chinese Han population. Eur J Hum Genet 19: 721-723.
- Njajou OT, Blackburn EH, Pawlikowska L, Mangino M, Damcott CM. et al. (2010) A common variant in the telomerase RNA component is associated with short telomere length. PLoS One 5: e13048.
- Codd V, Mangino M, van der Harst P, Braund PS, Kaiser M, et al. (2010) Common variants near TERC are associated with mean telomere length. Nat Genet 42: 197-199.
- Zee RY, Ridker PM, Chasman DI (2011) Genetic variants in eleven telomere-associated genes and the risk of incident cardio/cerebrovascular disease: The Women's Genome Health Study. Clin Chim Acta 412: 199-202.
- Heston LL, Mastri AR, Anderson VE, White J (1981) Dementia of the Alzheimer type. Clinical genetics, natural history, and associated conditions. Arch Gen Psychiatry 38: 1085-1090.
- Hofman A, Ott A, Breteler MM, Bots ML, Slooter AJ, et al (1997) Atherosclerosis, apolipoprotein E, and the prevalence of dementia and Alzheimer’s disease in a population-based study: The Rotterdam Study. Lancet 349: 151-154.
- Avramopoulos D, Mikkelsen M, Vassilopoulos D, Grigoriadou M, Petersen MB (1996) Apolipoprotein E allele distribution in parents of Down’s syndrome children. Lancet 347: 862-865.
- Petersen MB, Karadima G, Samaritaki M, Avramopoulos D, Vassilopoulos D, et al. (2000) Association between presenilin-1 polymorphism and maternal meiosis II errors in Down syndrome. Am J Med Genet 93: 366-372.
- Bhaumik P, Ghosh P, Ghosh S, Majumdar K, Pal S, et al. (2014) A Rare Intronic Variation of Presenilin-1 (rs201992645) is Associated with Alzheimer’s Disease and Down Syndrome Birth. Hereditary Genetics 3: 136.
- Potter H (1991) Review and hypothesis: Alzheimer disease and Down syndrome—chromosome 21 NDJ may underlie both disorders. Am J Hum Genet 48: 1192-1200.
- Hultén MA, Patel S, Jonasson J, Iwarsson E (2010) On the origin of the maternal age effect in trisomy 21 Down syndrome: the Oocyte Mosaicism Selection (OMS) model. Reproduction 139: 1-9.
- Ghosh S, Hong CS, Feingold E, Ghosh P, Ghosh P, et al. (2011) Epidemiology of Down syndrome: new insight into the multidimensional interactions among genetic and environmental risk factors in the oocyte. Am J Epidemiol 174: 1009-1016.