Review Article
Missegregation of Chromosome 21 In Oocyte: What Genetic Causes Imperil a Healthy Egg toHave Bad Fate?
Papiya Ghosh1 and Sujay Ghosh2*
Corresponding author: Dr. Sujay Ghosh, Department of Zoology, University of Calcutta, India,; E-mail: g_sujoy@yahoo.com
Citation: Ghosh P, Ghosh S. Missegregation of Chromosome 21 In Oocyte: What Genetic Causes Imperil a Healthy Egg to Have Bad Fate? J Cell Sci Molecul Biol. 2014;1(2): 109.
Copyright © 2014 Sujay Ghosh et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Cell Science & Molecular Biology | Volume: 1, Issue: 2
Reviewed & Approved by Dr. Surajit Sarkar, Assistant Professor, Department of Genetics, University of Delhi, India
Abstract
Down syndrome represents the most frequent live born aneuploidy and genetic form of intellectual disability. The overwhelming majority of live born Down syndrome is caused by trisomy 21 condition, although a small fraction is due to the inheritance of de novo translocated chromosome 21. The extra copy of chromosome 21 originates owing to non-separation or nondisjunction of chromosome at anaphase in meiotic cell division of gametogenesis. Sincereresearch attempt have recognized that the higher incidence of Down syndrome birth is associated with maternal advanced age of conception and reduced recombination on chromosome 21. Using panel of short tandem repeat (STR) markers scientists have proved for overwhelming majority of cases, the error originates in maternal first meiotic division when the oocyte grows in fetal ovary and the maternal age effect is restricted only to these maternally originated cases. Several hypotheses have been proposed to explain this maternal age associated increase in the incidence of Down syndrome birth. With large population sample and refined analytical approaches scientists have determined that the effect of recombination error on the nondisjunction is bimodal. On one hand, overall reduction in recombination frequency imparts a risk of nondisjunction irrespective of maternal age. On other hand, some susceptible chiasma configurations increase the chance of chromosome malsegregation with advancing maternal age. Thus the risk factors for the chromosome 21 nondisjunction are of two categories namely, maternal age independent and maternal age dependent. Beside these, some genetic polymorphisms showhigh degree of susceptibility for Down syndrome conception among women. The gene MTHFR is such candidate which is actually involved in folic acid metabolism pathway and its specific polymorphisms exhibits predisposition to missegregation of Ch21 irrespective of ethnicity of population across the globe. The present review is focused to address the latest development in understanding the role of genetic and molecular risk factors for Down syndrome birth.
Keywords: Down syndrome; Nondisjunction; Maternal Age; Recombination; Genetic polymorphism
Introduction
Down syndrome is the most frequent among all known recognizable live born human aneuploidies, estimatingapproximately 1 in 700 live-births [1] and for overwhelming cases it is caused by trisomy 21 chromosome complement. The trisomy 21 condition arises due to non-separation or nondisjunction (NDJ) of chromosome 21(Ch21) during gametogenesis and as a result disomic gametes form with two copies of a particular chromosome and upon dsfertilization by haploid gamete from opposite sex the trisomic fetus is formed. Beside chromosomal NDJ, a small proportion of DS occurs due to post zygotic mitotic error or translocation of chromosome 21 to other autosomes. Within the category of free trisomy 21 due to NDJ, overwhelming majority of errors occur in maternal oogenesis particularly at meiosis I (MI) stage. A little fraction of NDJ errors arise at paternal spermatogenesis.
In search of etiology of Ch21 NDJ, researchers have identified two risk factors unambiguously namely, advancing maternal age and altered pattern of meiotic recombination. Beside these two risk factors, other environmental and behavioural factors have also beenidentified associated with Ch21 NDJ, and they exhibit several degrees of interactions with advancing maternal age and recombination pattern of Ch21. These make the etiology of DS birth a puzzle in the field of medical genetics.
Advanced maternal age is the primary risk
The age of the mother at the time of the conception of a fetus with DS is, by far, the most significant risk factor for meiotic NDJ of Ch21. As a woman ages, her risk for having a fetus with trisomy 21 significantly increases. This association was noted initially by Penrose in 1933 [2]. For all the populations studied so far, estimated mean maternal age of conception of DS baby is higher than that of controls having euploid baby and women with MII NDJ is older than women affected with MI NDJ.
Hypotheses have been proposed to explain the relationship between advancing maternal age and higher incidence of aneuploid oocyte formation but no one has proved to be completely satisfactory. The most popular hypothesis [3] holds that the protracted tenure of oogenesis interrupted with meiotic halts (Figure 1), probably makes the eggs more vulnerable to the age effect than sperms. This preferential occurrence of maternal meiotic error is probably due to the mechanism of oocyte maturation in the ovary. Meiosis is initiated in the human foetal ovary at 11-12 weeks of gestation [3], but becomes arrested after completion of homologous chromosome pairing and recombination. This meiotic-halt lasts for several years until the elevated level of LH and FSH resume the process at the onset of puberty. Then the oocyte completes meiosis I (MI) and enters meiosis II (MII) and again undergoes a phase of pause. It completes the meiosis II after fertilization. Thus, the oocyte, whose ovulation marks the menarche, remains in pause for shortest period and the one that ovulates just preceding menopause experiences longest period of arrest. This long tenure of oocyte development makes it vulnerable to acquire environmental hazards within its microenvironment which inevitably increases the risk of chromosomal NDJ. The deterioration in egg includes diminishing amount of a meiotic proteins, like those maintaining sister chromatid adhesion [4,5] or meiotic checkpoints components [6] or weakening of centromere cohesion due to agerelated reduction in centromere associated proteins MCAK [7]. This list also includes accumulation of environmentally induced damage to the meiotic machinery over time or genetic changes such as mitochondrial deletions [8]. Among all these variables, the spindle assembly check point (SAC) components and sister chromatid cohesion (SCC) were investigated thoroughly [9], as they are prospective genetic candidate that may explain the maternal age effect on aneuploid oocyte formation. The SAC is a molecular machine that ensures proper chromosome separation in both mitosis and meiosis. In meiosis SAC prevents anaphase until all chromosomes properly attached to the spindle. The SAC includes MAD2L1, BUB1B, and TTK [10,11] which show decline in concentration with age in mouse leading to misaligned chromosomes [12] and suggests that errors in SAC function contribute to age-related aneuploidy. Disrupted spindles, misaligned chromosomes and decreased expression of SAC components Mad2L1 and Bub1 have evident in aged human oocytes [13,14] and the findings are consistent with hypothesis related to maternal aging. On the other hand, the SCC mediates physical pairing of duplicated chromosome which is essential for appropriate distribution of chromosomes. The cohesion along chromosome arms keeps bivalents intact in MI, and centromere cohesion holds sister chromatids together in MII. A defect in cohesion distal to crossover sites may result in a shift of chiasmata placement (alternatively known as ‘chiasma slippage’) or even premature bivalent separation in MI, whereas reduced centromere cohesion may result in premature separation of sister chromatids in MII [14]. The loss of cohesion with maternal age for distally placed chiasma [15] is consistent with the idea that cohesion defects may contribute to age related aneuploidy [9].
Another component that supposed to decline with age andcontributes significantly to maternal age effect on DS birth is the surveillance system of ovary that ensures achiasmate chromosome segregation [16]. Chiasma formation and subsequent recombination is prerequisite of faithful separation of homologues at meiotic anaphase. Absences of chiasma, faulty configurations of chiasma and reduction in chiasma frequency have been attributed; risk of NDJ of Ch21 and subsequent DS birth [17-19]. A high proportion of achiasmate Ch21 tetrad was reported among the mothers of DS having age >35 year [16]. As the decision regarding chiasma formation is taken in foetal ovary, high frequency of achisamate nondisjoined Ch21 in older oocyte can only be explained by down regulation of surveillance system. Human proteins involved in segregation of nonexchange chromosome show down regulation with increasing ovarian age [14,20].
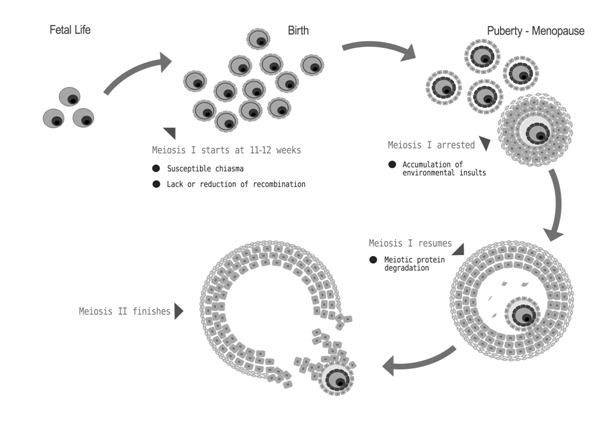
Figure 1: Phases of oocyte development and its various challenges that increases the risk of chromosomal nondisjunction.
A second hypothesis relates the “biological aging” or “ovarian aging” with the increasing rate of meiotic errors [21,22]. The central theme of this hypothesis is the prediction that biological aging is different among women of the same chronological age, and that the frequency of trisomic conceptions depend upon the biological age of the woman rather than the chronological age [21]. The biological age of women can usually be assessed by counting the falling number of antral follicles with chronological age together with decrease in total oocyte pool size [23,24]. These altogether alter the optimum hormonal balance in ovary, which is marked by falling concentration of serum inhibin A and B, decline in estrogens surge and elevated level of FSH [21]. This change in hormone balance is related to increased rate of aneuploidy at advanced maternal age. Support to this prediction is available from the experiments in mouse model [25]. Alternative to this prediction was provided in the ‘limited oocyte pool hypothesis’ [21] which stated that with biological age there is a decrease in thenumber of antral follicles, leaving only the premature or post mature oocyte to ovulate. The “biological aging” hypothesis predicts that women with a trisomic conception should on the average have an older “ovarian age” than other women of the same chronological age with a normal conception [21] and women having trisomic pregnancy have average earlier (~1 year) age of menopause [26]. If this were the case, one would expect that after a trisomic conception, the risk of a subsequent trisomy for any chromosome should be higher than the maternal age-related risk. In support to this prediction, recent data from prenatal diagnosis after a previous trisomic conception shows that the risk of a subsequent trisomy is about 1.7 times the maternal age-related risk [21]. Mathematical model proposed by Kline and Levin (1992) [27] estimated that women with trisomy pregnancy experience 0.9 years early menopause which suggests that such women suffer from advanced ovarian aging than the women with chromosomally normal pregnancies. Population sample survey for calculating the median age of menopause among the women with trisomic pregnancy loss also suggested an early cessation of menstrual cycle among them than the mothers with chromosomally normal foetus [26]. Elevated level of FSH is reported among the women with DS pregnancy [28,29] which suggests precocious aging among them. Very recently, Kline et al. (2011) [30] have conducted the survey on the hormonal level of women with trisomic pregnancy and supported the ‘reduced oocyte pool hypothesis’, suggesting that some women have smaller follicle content than the others of same chronological age. The former group is susceptible for rapid ovarian aging and associated trisomic conceptions. All these findings suggest intuitive existence of some predisposing factors among some women for their earlier aging that relates their trisomic conception too.
The third hypothesis is related to ‘genetic age’ of women and stated that it is the genetic aging of women that underlies all kind of degenerative changes in ovary and oocyte. The hypothesis was proposed by Ghosh et al., (2010) [18]. The authors estimated the telomere length of peripheral lymphocyte of women having DS child and compared the values with that of age matched controls. They found that beyond of age 29 years the DS bearing mothers exhibit rapid telomere attrition than do the controls. The authors inferred that DS bearing younger mothers do not experience any accelerated genetic aging; it is only the chronological older age when DS bearing mothers suffer from rapid genetic and molecular aging than do the age matched mothers of euploid child. The authors proposed ‘Genetic aging hypothesis’ which stated that some women are predisposed torapid genetic and molecular aging and its effect is exacerbated at advance age when age-related deteriorative changes also affects the chromosome separation system leading to NDJ. The notion suggested some intuitive link between telomere maintenance system (i.e., system of molecular aging) and chromosome segregating apparatus at molecular level.
Altered pattern of recombination and its interaction with maternal age
Aside from maternal age, there is only factor that has been shown to associate increased susceptibility of maternal NDJ, namely altered recombination patterns. Warren et al. (1987) [31] provided the first evidence to suggest that a proportion of maternal NDJ errors were associated with reduced recombination along Ch 21. Further examination has shown that, in addition to the absence of an exchange along the nondisjoined Ch 21, the placement of an exchange is an important susceptibility factor NDJ. Examination of recombination along the maternal nondisjoined Ch 21 has suggested three susceptible exchange patterns: 1) no exchange leads to an increased risk of MI errors, 2) a single telomeric exchange leads to an increased risk of MI errors, and 3) a pericentromeric exchange leads to an increased risk of so-called MII errors. These patterns are similar to those observed in model organisms where absent or reduced levels of recombination, along with sub-optimally placed recombinant events, increases the likelihood of NDJ [32-38]. Exchanges too close to the centromere or single exchange too close to the telomere seem to confer chromosomal instability.
Subsequently, researchers have identified a potential interaction between maternal age and pattern of recombination. The study on US population [39] provided the first evidence in this regard and proved an age related reduction in recombination frequency among the MI cases, with older women (35 yrs. and more) having less recombination along 21q than younger women (< 35 yrs.), as indicated by estimated length (cM) of age-specific linkage map of Ch21. In exploring the interaction between maternal age and recombination to gain further insight into the potential mechanisms of abnormal chromosome segregation, comparison had been performed [17] among women of various maternal ages who had an infant with DS due to a maternal MI error for frequency and location of meiotic exchanges along 21q. While there was no significant association between maternal age and overall frequency of exchange, the placement of meiotic exchange differed significantly by age of conception. In particular, single telomeric recombination event was present in highest proportion among the youngest age group (80%), while the proportion in the oldest group of women and in control group were almost equal (14% and 10% respectively). Moreover, studies [17,40] suggested that in maternal MI error cases, majority of single exchanges were located in the telomeric end of Ch21, whereas the single exchange within the peri-centromeric region was associated with maternal MII errors. In the independent age-stratified analysis on the US population by Oliver et al., (2008) [16] and on the Indian population by Ghosh et al., (2009) [19] a universal pattern of interactions among maternal age groups, chiasma placement and amount of meiotic recombination has been discovered. In these studies a major fraction of MI error was recorded due to absence of any detectable exchange between non-sister chromatids of nondisjoined homologues. A trend of decreasing frequency of achiasmate meiosis (meiosis without recombination) with increasing maternal age is also observed in both the studies [16,19], which suggests achiasmate meiosis without any recombination is maternal age-independent risk. According to the model of maternal risk factors for DS birth proposed by Oliver et al., (2008) [16] and supported by Ghosh et al. (2009, 2010) [18,19] that any risk factor which is maternal age independent should present in highest frequency in the younger mother, the age group in which other risk factors are usually absent. In contrast, any risk factorswhose frequency increases with increasing maternal age is regarded as maternal age dependent risk factor as its effect gets exacerbated in interaction with increasing maternal age. The chiasma stabilizes the tetrad and counter balances the pull from opposite poles which ensure the faithful segregation of homologues. In absence of chiasma, the chromosomes move randomly at MI, resulting in formation of disomic gametes. As the chiasma formation takes place in foetal ovary, the achisamate chromosome containing disomic oocyte may ovulate at any time in reproductive life and hence it is maternal age independent risk factor of Ch21 NDJ.
In both the studies on US and Indian populations [16,19] the single telomeric chiasma and subsequent recombination were found in highest frequency among the women of younger age group i.e., age group below 29 years, who had NDJ error at meiosis I stage of oogenesis and there were a gradual decrease in telomeric chiasma frequency with advancing maternal age. This observation suggests that the single telomeric chiasma formation is the risk of NDJ of Ch 21 even in younger women who otherwise do not suffer from deterioration related to the aging. Thus within the total risk probability of Ch21 NDJ, the single telomeric chiasma formation represent the highest proportion among the younger women of MI NDJ category. Two important inferences have been drawn from this finding. The first one is that the single telomeric chiasma formation is maternal age independent risk of Ch21 NDJ. The second is that the single telomeric chiasma probably induces some structural instability of Ch21 that segregates randomly at meiosis I which takes place in fetal ovary.
Understanding of the mechanism how single telomeric chiasma causes chromosomal mis-segregation has been obtained from the observation in model organisms like Drosophila [36], Saccharomyces [37] and Caenorhabditis elegans [35]. As the telomeric chiasma located far from the kinetochore, the point of spindle-attachment links the homologues less efficiently and orients each kinetochore to the same spindle pole and prevents bi-orientation of homologues [41,42,36]. Most likely, this susceptibility is related to the minimal amount of sister chromatid cohesion complex remaining distal to the exchange event [43]. Alternatively, the integrity of chiasma may be compromised when a minimum amount of cohesin remains to hold homologue together. Thus bivalent may act as pair of functional univalent during MI, as has been evident in human oocyte [44,455].
Another chiasma configuration that poses susceptibility for NDJ of Ch21 is the pericentromeric exchange. In both the studies on US and Indian DS populations [16,19], highest frequency of pericentromeric exchange was scored in older women having age >34 years. A trend of gradual increase in centromeric chiasma frequency with increasing age was recorded in both the studies with gradual shifting of chiasma from middle of the chromosome in younger age group to more proximal to centromere in older age group.
In explaining the effect on chromosome segregation that single centromeric chiasma imparts two hypotheses have been put forward by the authors. Chiasma that is positioned very close to centromere may cause ‘chromosomal entanglement’ at MI, with the bivalent being unable to separate, passing intact to MII metaphase plate [40]. Upon MII division, the bivalent divides reductionally, resulting in disomic gamete with identical centromeres. In this manner, proximal pericentromeric exchange, which occurs at MI, is resolved and visualized as MII error. According to an alternate model, studied in Drosophila [36], proximal chiasmata lead to a premature sister chromatid separation just prior to anaphase I. Resolution of chiasmata requires the release of sister chromatid cohesion distal to the site of exchange [42]. Attempt to resolve chiasmata that are very near to centromere could result in premature separation of chromatids. If the sister chromatids migrate to a common pole at MI, they have 50% probability to move randomly into the same product of meiosis at MII, resulting in an apparent MII NDJ. Similar observation is reported from the study in Yeast in which centromereproximal crossover promotes local loss of sister-chromatid cohesion [46]. Studies of NDJ in both human [45] and Drosophila [47] have provided preliminary supports for this model.
The effect of pericentromeric exchange on meiotic chromosome separation gets exacerbated with maternal age related insults in ovarian environment, as indicated by greater proportion of DS births among older women who have experienced the particular pattern of chiasma formation. This relationship can be interpreted in two different ways: 1) pericentromeric exchange set up a suboptimal configuration that initiates or exacerbates the susceptibility to maternal age-related risk factors, perhaps leading to an increase in premature sister chromatid segregation or 2) a pericentromeric exchange protect the bivalent against age related risk factor, allowing proper segregation of homologues, but not the sister chromatids at MII [16]. The former explanation is likely to the ‘two hit model’ proposed previously by Lamb et al., (1996) [40]. Alternatively, a pericentromeric exchange may protect the bivalent from maternal age related risk factors. The effect of degradation of centromere or sister chromatid cohesion complexes or of spindle proteins with age of oocyte may lead to premature sister chromatid separation. Perhaps the pericentromeric exchanges help to stabilize the compromised tetrad through MI. This would lead to an enrichment of MII errors among the older oocytes which is a maternal age dependent risk for NDJ of Ch21.
As far as effect of multiple chiasmata formation on thenondisjoined Ch 21 is concerned, two important reports havebeen published very recently. In their study Ghosh et al. (2010) [19] found that two or more chiasmata formation is prevalent particularly in older age group (≥ 34 years). This signifies that even when Ch21 experiences formation of two or more chiasmata which are believed to be protective of NDJ, particularly when aging effect imparts various degenerative changes in ovary. Analyzing the effect of multiple chiasmata of the 21q, Oliver et al. (2011) [48] found a decrease in the interval between two simultaneous chisamata on the chromosome that disjoined at MI and this closeness is due to shifting of distal chiasma towards centromere. The author argued that as the proximal chiasma remains at its usual position, similar to that on the normally disjoined chromosome, it is the distal chiasma whose dislocation towards the proximal chiasma nullifies the ‘good-effect’ of the latter that is needed for faithful segregation of the chromosome. The Ch21 experiences such distal chiasma dislocation in association with correctly placed proximal chiasma disjoined erroneously at MI. Moreover, the authors found more intimate positioning of proximal chiasma with the centromere of the chromosomes with two exchange and this tendency increases with advancing age. This pattern is very similar to the single chiasma shifting related to MII errors reported in earlier studies [16,19]. Moreover, the authors further extend their realization that the centromeric chiasma may not be protective of NDJ, the notion previously assumed both by Oliver et al. (2008) [16] and Ghosh et al. (2009) [19].
Very recently, a denser recombination map along the long arm of nondisjoined Ch21 has been made using SNP (single nucleotide polymorphism) markers by Oliver et al. (2014) [49]. The authors enquire whether the altered patterns of recombination associated with maternal nondisjunction of Ch 21 could be explained by differences in the relationship between recombination placement and recombination-related genomic features (i.e., GC content, Chg. fraction, Poly(A)/Poly(T) fraction or gene density) on 21q or differential hot-spot usage along the no disjoined chromosome 21 (Oliver et al. 2014). They included a total of 297 maternal MI and 277 maternal MII cases of trisomy 21 and genotyped them at 1536 SNP loci on 21q using the Illumine Golden Gate Assay platform. Authors have identified several recombination hot-spots along the length of 21q and examined proportion of single and double recombination events within these hotspots and compared them with the pattern of hot-spot use among controls. For MI cases, the use of recombination hotspot did not differ significantly from the control, which suggests altered pattern of recombination among MI cases was not due to differential use of hot-spots. For MII single recombinant events compared with those for MI-single recombinants events and controls, authors found proportion of single recombinant within the hot spot is significantly positively correlated. The authors infer that potentiallyfactors characteristic of pericentromeric DNA such as chromatin structure or epigenetic modifications may affect the accessibility of a specific chromosome region to recombination in at least a proportion of oocytes with meiotic errors. In analysis of double recombinantsevents, the authors found similar results with respect defined hotspots among MI and MII errors. They detected a significant relationship between defined hotspots for the distal recombinant events among doubles, but not the proximal events. Furthermore, the lack of evidence for an association in the proximal region differed from that in controls where an association was detected (i.e., significant interaction). The authors did not provide consistent evidence that genomic features present at the site of recombination or differential hotspot usage are implicated in the nondisjunction of Ch 21.
Genetic polymorphism and increasing susceptibility of Down syndrome birth
Maternal genetic factors such as polymorphisms of certain genes probably make them susceptible for NDJ error. Experimental organisms have been used to identify genes that are important in theproper segregation of chromosomes. The potential candidates are those genes involved in the meiotic process such as homolog pairing, assembly of the synaptonemal complex, chiasmata formation and chiasma positioning, sister chromatid cohesion, spindle formation. Genetic variations of these genes are predisposing factors for chromosome NDJ. Study on Yeast revealed that synaptonemal complex protein Zip1 is essential for nonexchange chromosome segregation and mutation of it causes NDJ [50]. Mutation in synaptonemal complex protein SYP3 is known to associate the recurrent pregnancy loss due to abnormal chromosome behavior [51]. Yeast med1 mutant revealed both meiotic homologous NDJ and precocious separation of sister chromatids [52]. In Drosophila the SAC component BubR1 has been proved to be essential for sister chromatid cohesion and synaptonemal complex and mutation of this gene causes NDJ [53]. Unfortunately, mutation or polymorphisms of these genes have not been examined among the women bearing DS child.
The gene that has been identified first in this category is MTHFR (methylene tetrahydrofolate reductase), which is not directly related to the meiotic process. The case-control study by James et al., (1999) [54] provided preliminary evidence that the 677C→ T polymorphism in the MTHFR gene increases the risk of having a child with DS (Odds Ratio = 2.6) in North American population. This polymorphism is associated with elevated plasma homocysteine and/or low folate status [55]. Folate is essential for the production of S-adenosylmethionine, which is the primary methyl donor (Figure 1) for epigenetic DNA methylation and essential for gene expression regulation and maintenance of chromosomal integrity at centromere [54,56,57]. Folate deficiency reduces S-adenosylmethionine synthesis, leading to DNA hypomethylation [58-60]. The pericentromeric hypomethylation could impair the heterochromatin formation and kinetochore establishment (Figure 2) resulting in chromosomal NDJ [54]. This can happen because the stable centromeric chromatin depends on the epigenetic inheritance of specific centromeric methylation patterns and it binds with specific methyl-sensitive proteins in order to maintain the higher-order DNA architecture necessary for kinetochore assembly [61].
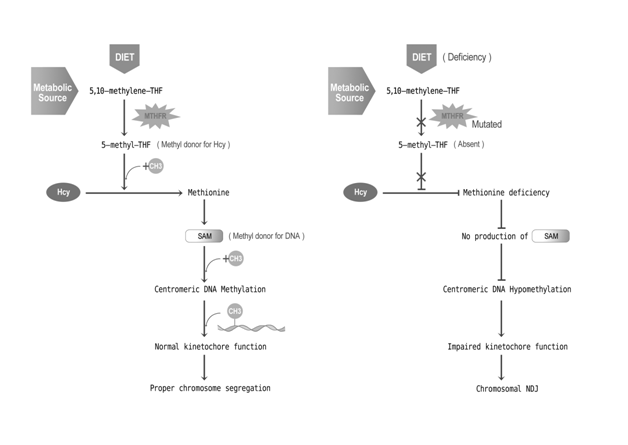
Figure 2: Pathway for folic acid metabolism and mutation in MTHFR gene that increases the risk of chromosomal nondisjunction.
This initial report had inspired several follow-up studies on the MTHFR 677C→ T polymorphism, as well as several other allelic variants in the folate pathway genes to identify genetic risk factors for having a child with DS. But the results are inconsistent [62,63]. Those who have examined blood homocysteine levels, a broad-spectrum indicator of nutritional and/or genetic impairment in folate/B12 metabolism have documented a significantly higher level among the mothers of children with DS compared with control mothers from the same country. One possible explanation for the inconsistent results among the numerous studies may reflect the complex interaction between effects of genetic variants and nutritional intake[63]. Nevertheless, support to the notion regarding the association between MTHFR 677C-T polymorphism and risk of DS birth was provided by other studies in different populations. Wang et al., (2004) [60] reported significant increase in the risk of DS conception among Chinese women bearing two polymorphisms namely, polymorphisms of MTHFR 677C→ T and the polymorphism MTRR (Methionine synthase reductase) 66A→ G. The estimated risks were more than three folds and five folds for MTHFR (Odd Ratio=3.7; 95% CI, 1.78~8.47) and MTRR (Odd Ratio= 5.2; 95% CI, 1.90~14.22) respectively. The combined presence of both polymorphisms was associated with a greater risk of DS than the presence of either alone, with an odds ratio of 6.0 (95% CI, 2.058~17.496). The study on Italian population also agreed the link between DS birth and MTHFR and MTRR polymorphisms [64,65/a>]. Cyril et al., (2009) conducted such association study on Indian women and confirmed the association of MTHFR 677C→ T polymorphism with DS birth risk.
The other way to find out the genes involved in human NDJ is to analyze the association of consanguinity and trisomy 21 [55]. If such an association really does exist, it would provide evidence for a genetic effect for NDJ. The study of Alfi et al., (1980) [66] provided one of the earlier reports suggesting an association between increased consanguinity among parents of individuals with DS in a study population in Kuwait. Authors postulated the existence of a gene that increases the risk for mitotic NDJ. Alternatively, they suggested that increased rates of consanguinity among parents would be correlated with those in grandparents and therefore, an autosomal recessive gene may be postulated to be involved in meiotic NDJ in the homozygous parents. But the reports from subsequent studies in other populations are contradictory and did not find any evidence for an association between consanguinity and human NDJ [67-72].
Lastly, differences in the prevalence of DS among different racial groups may provide indirect evidence for genetic factors involved in human NDJ. However, such studies are difficult to conduct and to interpret. Differences (or similarities) may reflect the maternal age distribution of the population, accuracy of diagnosis, cultural preference and/or access to selective prenatal termination of pregnancies with trisomic fetuses, and as yet unidentifiedenvironmental factors [55]. Only one such study by Allen et al., (2009) [73] reported demographic differences in mean maternal age of DS conception recorded in two different sample sets from USA. This study included DS samples from Atlanta Down syndrome project and National Down syndrome project and found that mothers enrolled in National Down syndrome project are on an average older than those of Atlanta. Moreover, the authors have also reported some ethnic differences in maternal age distribution. The Atlanta Down syndrome project had a higher proportion of cases and controls that were black and a significantly smaller proportion of Hispanics than did the National Down syndrome project. Comparison of mean maternal ages indicated variation by ethnic groups. In both the Atlanta Down syndrome project and National Down syndrome project, white mothers tended to be older than their black or Hispanic counterparts. Specifically, for both cases and controls, white mothers were found to be significantly older than black mothers (P< 0.01) and Hispanic mothers (P< 0.01); blacks and Hispanics were not significantly different from each other (P>0.05). To confirm such effect of demographic and ethnic differences on the etiology of DS birth large scale population based studies are needed to be conducted.
Future Research
Attempt to resolve the etiology of DS birth is a continuous process and we hope this will bring new insight in the understanding the hidden truth in near future. But the problem lies in its multi factorial nature, which inevitably suggests necessity of multi-faceted research efforts from the several directions. For example, it is needed to analyze the polymorphisms of certain genes that regulate meiotic recombination or genes that control maternal molecular aging or those who are involved in faithful chromosome segregation system in meiosis. For searching the cause of recombination anomaly, PRDM9 would be the good target of investigation, as it is a documented regulator of mammalian recombination [74,75]. Telomere maintenance system and their genetic components such as TERT and TERC may be the other targets of research and would help us to realize the cause of molecular aging and related genetic susceptibility of NDJ [76,77]. The component of sister chromatid cohesion complex and their role in chromosome segregation have been evident in mammals and nonmammalian model organisms. Their functional impairment is known to be associated with elevated level of chromosomal missegregation and aneuploidy [78-80]. But their role and allelic variation have not been explored in the context of Ch21 NDJ and subsequent DS birth. Apart from genetic components, several environmental influences are known to associate with DS birth as risk factors. But proper molecular study on how their adverse effect interacts and imperils faithful chromosome separation apparatus is tantalizingly low. At this level it is almost certain that environmental hazards or aneugen in various forms is associated with accidental increase in DS birth rateat different parts of world. But scientific evidence in favor of their interaction with genetic component is lacking and needs indepth study. If these could be resolved properly in future great advances will be made in the field of medical science and potential couple would enjoy their parenthood with physically and mentally healthy babies.
Acknowledgements
We are extremely grateful to Prof. Eleanor Feingold, Pittsburgh University, USA and Prof. Stephanie Sherman, Emory University, Atlanta, USA for their cooperation in Down syndrome research.
References
- Kanamori G, Witter M, Brown J, Williams-Smith L (2000) Otolaryngolog manifestations of Down Syndrome. Otolaryngol Clin North Am 33: 1285-1292.
- Penrose LS (1933) The relative effect of paternal and maternal age in Mongolism. J Genet 27: 219-224.
- Gondos B, Westergaard L, Byskov AG (1986) Initiation of oogenesis in the human fetal ovary: ultrastructural and squash preparation study. Am J Obstet Gynecol 155: 189-195.
- Hodges CA, Revenkova E, Jessberger R, Hassold TJ, Hunt PA (2005) SMC1beta-deficient female mice provide evidence that cohesins are a missing link in age-related nondisjunction. Nat Genet 3: 1351-1355.
- Hunt PA, Hassold TJ (2008) Human female meiosis: What make a good egg go bad? Trend Genet 24: 86-93..
- Garcia-Cruz R, Brieno MA, Roig I, Grossmann M, Velilla E, et al. (2010) Dynamics of cohesin proteins REC8, STAG3, SMC1 beta and SMC3 are consistent with a role in sister chromatid cohesion during meiosis in human oocytes. Hum Reprod 25: 2316-2327.
- Eichenlaub-Ritter U, Staubach N, Trapphoff T (2010) Chromosomal and cytoplasmic context determines predisposition to maternal age-related aneuploidy: brief overview and update on MCAK in mammalian oocytes. Biochem Soc Trans 38: 1681-1686.
- Van Blerkom J (2011) Mitochondrial function in the human oocyte and embryo and their role in developmental competence. Mitochondrion 11: 797-813.
- Chiang T, Duncan FE, Schindler K (2010) Evidence that weakened centromere cohesion is a leading cause of age-related aneuploidy in oocytes. Curr Biol 20: 1522-1528.
- Hached K, Xie SZ, Buffin E, Cladière D, Rachez C (2011) Mps1 at kinetochores is essential for female mouse meiosis I. Development 138: 2261-2271.
- Niault T, Hached K, Sotillo R, Sorger PK, Maro B, et al. (2007) Changing Mad2 levels affects chromosome segregation and spindle assembly checkpoint control in female mouse meiosis I. PLoS One 2: e1165.
- Pan H, Ma P, Zhu W, Schultz RM (2008) Age-associated increase in aneuploidy and changes in gene expression in mouse eggs. Dev Biol 316: 397-407.
- McGuinness BE, Anger M, Kouznetsova A, Gil-Bernabé AM, Helmhart W, et al. (2009). Regulation of APC/C activity in oocytes by a Bub1-dependent spindle assembly checkpoint. Curr Biol 19: 369-380.
- Steuerwald N, Cohen J, Herrera RJ, Sandalinas M, Brenner CA (2001) Association between spindle assembly checkpoint expression and maternal age in human oocytes. Mol Hum Reprod 7: 49-55.
- Subramanian VV, Bickel SE (2008) Aging predisposes oocytes to meiotic nondisjunction when the cohesin subunit SMC1 is reduced. PLoS Genet 4: e1000263.
- Oliver TR, Feingold E, Yu K, Cheung V, Tinker S, et al. (2008) New insights into human nondisjunction of chromosome 21 in oocytes. PLoS Genet 4: e1000033.
- Lamb NE, Sherman SL, Hassold TJ (2005) Effect of meiotic recombination on the production of aneuploid gametes in humans. Cytogenet Genome Res 111: 250-255.
- Ghosh S, Feingold E, Chakraborty S, Dey SK (2010) Telomere length is associated with types of chromosome 21 nondisjunction: a new insight into the maternal age effect on Down syndrome birth. Hum Genet 127: 403-409.
- Ghosh S, Feingold E, Dey SK (2009) Etiology of Down Syndrome: Evidence for Consistent Association among Altered Meiotic Recombination, Nondisjunction and Maternal Age Across Populations. Am J Med Genet 149A: 1415-1420.
- Baker DJ, Jeganathan KB, Cameron JD, Thompson M, Juneja S, et al. (2004). BubR1 insufficiency causes early onset of aging associated phenotypes and infertility in mice. Nat Genet 36: 744-749.
- Warburton D (1989) The effect of maternal age on the frequency of trisomy: change in meiosis or in utero selection? Prog Clin Biol Res 31: 165-181.
- Warburton D (2005) Biological aging and etiology of aneuploidy. Cytogenetics and Genome Res 111: 266-272.
- Scheffer GJ, Broekmans FJ, Dorland M, Habbema JD, Looman CW, et al. (1999) Antral follicle counts by transvaginal ultrasonography are related to age in women with proven fertility. Fertil Steril 72: 845-851.
- Kline J, Kinney A, Reuss ML, Kelly A, Levin B, et al. (2004) Trisomic pregnancy and the oocyte pool. Hum Reprod 19: 1633-1643.
- Roberts R, Iatropoulou A, Ciantar D, Stark J, Becker DL, et al. (2005) Follicle-stimulating hormone affects metaphase I chromosome alignment and aneuploidy in mouse oocytes matured in vitro. Biol Reprod 72: 107-118.
- Kline J, Kinney A, Levin B, Warburton D (2000) Trisomic pregnancy and earlier age at menopause. Am J Hum Genet 67: 395-404.
- Kline J, Levin B (1992) Trisomy and age at menopause: predicted associations given a link with rate of oocyte atresia. Pediatr Perinat Epidemiol 6: 225-239.
- Nasseri A, Mukherjee T, Grifo JA, Noyes N, Krey L, et al. (1999) Elevated day 3 serum follicle stimulating hormone and/or estradiol may predict fetal aneuploidy. Fertil Steril 71: 715-718.
- van Montfrans JM, van Hooff MH, Martens F, Lambalk CB (2002) Basal FSH, estradiol and inhibin B concentrations in women with a previous Down’s syndrome affected pregnancy. Hum Reprod 17: 44-47.
- Kline JK1, Kinney AM, Levin B, Kelly AC, Ferin M, et al. (2011) Trisomic pregnancy and elevated FSH: implications for the oocyte pool hypothesis. Hum Reprod 26 :1537-1550.
- Warren AC, Chakravarti A, Wong C, Slaugenhaupt SA, Halloran SL, et al. (1987) Evidence for reduced recombination on the nondisjoined chromosomes 21 in Down syndrome. Science 237: 652-654.
- Rasooly RS, New CM, Zhang P, Hawley RS, Baker BS (1991) The lethal(1)TW-6cs mutation of Drosophila melanogaster is a dominant antimorphic allele of nod and is associated with a single base change in the putative ATP-binding domain. Genetics 129: 409-422.
- Moore DP, Miyazaki WY, Tomkiel JE, Orr-Weaver TL (1994) Double or nothing: a Drosophila mutation affecting meiotic chromosome segregation in both females and males. Genetics 136: 953-964.
- Sears DD, Hegemann JH, Shero JH, Hieter P (1995) Cis-acting determinants affecting centromere function, sister-chromatid cohesion and reciprocal recombination during meiosis in Saccharomyces cerevisiae. Genetics 139: 1159-1173.
- Zetka M, Rose A (1995) The genetics of meiosis in Caenorhabditis elegans. Trends Genet 11: 27-31.
- Koehler KE, Hawley RS, Sherman S, Hassold T (1996) Recombination and nondisjunction in humans and flies. Hum Mol Genet 5: 1495-1504.
- Ross LO, Maxfield R, Dawson D (1996) Exchanges are not equally able to enhance meiotic chromosome segregation in yeast. Proc Natl Acad Sci USA 93: 4979-4983.
- Krawchuk MD, Wahls WP (1999) Centromere mapping functions for aneuploid meiotic products: Analysis of rec8, rec10 and rec11 mutants of the fission yeast Schizosaccharomyces pombe. Genetic 153: 49-55.
- Sherman SL, Petersen MB, Freeman SB, Hersey J, Pettay D, et al. (1994) Non-disjunction of chromosome 21 in maternal meiosis I: evidence for a maternal age-dependent mechanism involving reduced recombination. Hum Mol Genet 3: 1529-1535.
- Lamb NE, Freeman SB, Savage-Austin A, Pettay D, Taft L, et al. (1996) Susceptible chiasmate configurations of chromosome 21 predispose to non-disjunction in both maternal meiosis I and meiosis II. Nat Genet 14: 400-405.
- Nicklas RB (1974) Chromosome segregation mechanisms. Genetics 78: 205-213.
- Hawley RS, Frazier JA, Rasooly R (1994) Separation anxiety: the etiology of nondisjunction in flies and people. Hum Mol Genet 3: 1521-1528.
- Orr-Weaver T (1996) Meiotic nondisjunction does the two-step. Nat Genet 14: 374-376.
- Angell R (1994) Aneuploidy in older women. Higher rates of aneuploidy in oocytes from older women. Hum Reprod 9: 1199-2000.
- Angell R (1995) Mechanism of chromosome nondisjunction in human oocytes. Prog Clin Biol Res 393: 13-26.
- Rockmill B, Voelkel-Meiman K, Roeder GS (2006) Centromere-proximal crossovers are associated with precocious separation of sister chromatids during meiosis in Saccharomyces cerevisiae. Genetics 174: 1745-1754.
- Miyazaki WY, Orr-Weaver TL (1992) Sister-chromatid misbehavior in Drosophila ord mutants. Genetics 132 : 1047-1061.
- Oliver TR, Tinker SW, Allen EG, Hollis N, Locke AE, et al. (2012) Altered patterns of multiple recombinant events are associated with nondisjunction of chromosome 21. Hum Genet 131: 1039-1046.
- Oliver TR, Middlebrooks CD, Tinker SW, Allen EG, Bean LJ, et al. (2014) An examination of the relationship between hotspots and recombination associated with chromosome 21 nondisjunction. PLoS One 9: e99560.
- Newnham L1, Jordan P, Rockmill B, Roeder GS, Hoffmann E (2010) The synaptonemal complex protein, Zip1, promotes the segregation of nonexchange chromosomes at meiosis I. Proc Natl Acad Sci U S A 107: 781-785.
- Bolor H1, Mori T, Nishiyama S, Ito Y, Hosoba E, et al. (2009) Mutations of the SYCP3 gene in women with recurrent pregnancy loss. Am J Hum Genet 84: 14-20.
- Rockmill B, Roeder GS (1994) The yeast med1 mutant undergoes both meiotic homolog nondisjunction and precocious separation of sister chromatids. Genetics 136: 65-74.
- Malmanche N1, Owen S, Gegick S, Steffensen S, Tomkiel JE, et al. (2007) Drosophila BubR1 is essential for meiotic sister-chromatid cohesion and maintenance of synaptonemal complex. Curr Biol 17: 1489-1497.
- James SJ, Pogribna M, Pofribny IP, Melnyk S, Hine RJ, et al. (1999) Abnormal folate metaboloism and mutation in the methylenetetrahydrofolate reductase gene may be maternal risk factors for Down syndrome. Am J Cli Nut 70: 495-501.
- Sherman SL, Freeman SB, Allen EG, Lamb NE (2005) Risk factors for nondisjunction of trisomy 21.Cytogenet Genome Res 111: 273-280.
- Dworkin AM, Tober KL, Duncan FJ, Yu L, VanBuskirk AM, et al. (2009) chromosomal aberrations in UVB-induced tumors of immunosuppressed mice. Genes Chromosomes Cancer 48: 490-501.
- Sciandrello G, Caradonna F, Mauro M, Barbata G (2004) Arsenic-induced DNA hypomethylation affects chromosomal instability in mammalian cells. Carcinogenesis 25: 413-417.
- Pogribny IP, Muskhelishvili L, Miller BJ, James SJ (1997) Presence and consequence of uracil in preneoplastic DNA from folate/methyl-deficient rats. Carcinogenesis 18: 2071-2076.
- Beetstra S, Thomas, P, Salisbury C, Turner J, Fenech M (2005) Folic acid deficiency increases chromosomal instability, chromosome 21 aneuploidy and sensitivity to radiation-induced micronuclei. Mutat Res 578: 317-326.
- Wang X, Thomas P, Xue J, Fenech M (2004) Folate deficiency induces aneuploidy in human lymphocytes in vitro - evidence using cytokinesis-blocked cells and probes specific for chromosomes 17 and 21. Mutat Res 551: 167-180.
- Migliore L, Migheli F, Coppedè F (2009) Susceptibility to aneuploidy in young mothers of Down syndrome children. Scientific World Journal 9: 1052-1060.
- James SJ (2004a) Maternal metabolic phenotype and risk of Down syndrome: beyond genetics. Am J Med Genet A 127: 1-4.
- James SJ (2004b) Response to letter: Down syndrome and folic acid deficiency. Am J Med Genet A 131, 328-329.
- Coppedè F, Grossi E, Migheli F, Migliore L (2010) Polymorphisms in folate-metabolizing genes, chromosome damage, and risk of Down syndrome in Italian women: identification of key factors using artificial neural networks. BMC Med Genomics 3: 42.
- Cyril C, Rai P, Chandra N, Gopinath PM, Satyamoorthy K (2009) MTHFR Gene variants C677T, A1298C and association with Down syndrome: A Case-control study from South India. Indian J Hum Genet 15 : 60-64.
- Alfi OS, Chang R, Azen SP (1980) Evidence for genetic control of nondisjunction in man. Am J Hum Genet 32: 477-483.
- Devoto M, Prosperi L, Bricarelli FD, Coviello DA, Croci G, et al. (1985) Frequency of consanguineous marriages among parents and grandparents of Down patients. Hum Genet 70: 256-258.
- Hamamy HA, al-Hakkak ZS, al-Taha S. (1990) Consanguinity and the genetic control of Down syndrome. Clin Genet 37: 24-29.
- Basaran N, Cenani A, Sayli BS, Ozkinay C, Artan S, et al. (1992) Consanguineous marriages among parents of Down patients. Clin Genet 42: 13-15.
- Zlotogora J (1997) Genetic disorders among Palestinian Arabs: 1. Effects of consanguinity. Am J Med Genet 68: 472-475.
- Sayee R, Thomas IM (1998) Consanguinity, non-disjunction, parental age and Down’s syndrome. J Indian Med Assoc 96: 335-337.
- Rittler M, Liascovich R, Lopez-Camelo J, Castilla EE (2001) Parental consanguinity in specific types of congenital anomalies. Am J Med Genet 102: 36-43.
- Allen EG, Freeman SB, Druschel C, Hobbs CA, O'Leary LA, et al. (2009). Maternal age and risk for trisomy 21 assessed by the origin of chromosome nondisjunction: a report from the Atlanta and National Down Syndrome Projects. Hum Genet 125: 41-52.
- Jeffreys AJ, Cotton VE, Neumann R, Lam KW (2013) Recombination regulator PRDM9 influences the instability of its own coding sequence in humans. Proc Natl Acad Sci U S A 110: 600-605.
- Berg IL, Neumann R, Sarbajna S, Odenthal-Hesse L, Butler NJ, Jeffreys AJ (2011) Variants of the protein PRDM9 differentially regulate a set of human meiotic recombination hotspots highly active in African populations. Proc Natl Acad Sci U S A 108: 12378-12383.
- Lee JH, Cheng R, Honig LS, Feitosa M, Kammerer CM, et al. (2014) Genome wide association and linkage analyses identified three loci-4q25, 17q23.2, and 10q11.21-associated with variation in leukocyte telomere length: the Long Life Family Study. Front Genet 4:310.
- Zhou J, Ding D, Wang M, Cong YS (2014) Telomerase reverse transcriptase in the regulation of gene expression. BMB Rep 47: 8-14.
- Vogt E, Kipp A, Eichenlaub-Ritter U (2009) Aurora kinase B, epigenetic state of centromeric heterochromatin and chiasma resolution in oocytes. Reprod Biomed Online 19: 352-368.
- Katis VL, Galova M, Rabitsch KP, Gregan J, Nasmyth K (2004) Maintenance of cohesin at centromeres after meiosis I in budding yeast requires a kinetochore-associated protein related to MEI-S332.Curr Biol 14: 560-572.
- Balicky EM, Endres MW, Lai C, Bickel SE (2002) Meiotic cohesion requires accumulation of ORD on chromosomes before condensation. Mol Biol Cell 13: 3890-3900.