Review Aricle
Neutrophil Gelatinase-Associated Lipocalin (NGAL): A Promising Biomarker for Cancer Diagnosis and A Potential Target for CancerTherapeutics
Ajaikumar B. Kunnumakkara*
Corresponding author: Ajaikumar B. Kunnumakkara, Cancer Biology Laboratory, Department of Biotechnology, Indian Institute of Technology Guwahati, Assam-781039, India, Phone : +91 361 258 2231 (Office), +91 789 600 5326 (Mobile), Fax: +91 361 258 2249 (Office); E-mail: kunnumakkara@iitg.ernet.in; ajai78@gmail.com
Citation: Monisha J, Padmavathi G, Bordoloi D, Roy NK, Kunnumakkara AB. Neutrophil Gelatinase-Associated Lipocalin (NGAL): A Promising Biomarker for Cancer Diagnosis and A Potential Target for Cancer Therapeutics. J Cell Sci Molecul Biol. 2014;1(2): 106.
Copyright © 2014 Ajaikumar B. Kunnumakkara et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Cell Science & Molecular Biology | Volume: 1, Issue: 2
Submission: 06/06/2014; Accepted: 27/06/2014; Published: 02/07/2014
Reviewed & Approved by Dr. Indranil Chattopadhyay, Assistant Professor, Department of Life Sciences, Central University of Tamil Nadu, India
Abstract
NGAL, also known as lipocalin-2, is a secretory glycoprotein encoded by LCN2 gene and found to be expressed majorly in kidney diseases. In recent years, NGAL is procuring enormous attention as a cancer biomarker with its differential expression pattern. In breast cancer, NGAL binds with MMP9 and promotes invasion. High expression of NGAL is strongly involved in the progression of colorectal cancer, esophageal cancer, chronic myeloid leukemia, hepatocellular carcinoma, gastric and endometrial cancers. However, in other cancers such as pancreatic cancer and oral cancer, downregulation of NGAL leads to disease progression. Interestingly, a cell surface receptor for NGAL, NGALR was identified and the co-expression of both NGAL and NGALR has been implicated in different cancers. Therefore, understanding the function of NGAL in the development of cancer would provide us new insights to cancer development. This would also help us to develop biomarkers for cancer diagnosis and prognosis. This review article discusses the role of NGAL and NGALR in malignant diseases.
Keywords: NGAL; NGALR; LCN2; Cancer; Biomarker; Invasion; Metastasis
Abbreviations
CML: Chronic Myeloid Leukemia; C/EBP: CCAAT-Enhancer- Binding Proteins; EGFR: Epidermal Growth Factor Receptor; ESCC: Esophageal Squamous Cell Carcinoma; iAS: Inorganic Arsenic; MMP9: Matrix Metalloproteinase 9; NF-κB: Nuclear FactorKappa B; NGAL: Neutrophil Gelatinase- associated Lipocalin; NGALR: Receptor NGAL; PaCa: Pancreatic Cancer; pNGAL: Plasma NGAL; STAT3- Signal Transducer and activator of Transcription 3; SV40: Simian Virus 40; TPA: 12-O tetradecanoylphorbol-13-acetate; TRE: TPA Responsive Element.
Introduction
The lipocalin protein family is a huge group of small secretedglycoproteins with functional diversity. Various functions of these proteins include retinol transport, cryptic coloration, olfaction, pheromone transport, enzymic synthesis of prostaglandins, regulation of the immune response and mediation of cell homoeostasis [1]. Members of this family share a common secondary and tertiary structure known as “the lipocalin fold”. The lipocalin fold is a barrel like structure, consisting of eight antiparallel beta sheets linked with hydrogen bonds and helps enormously in ligand binding. The connection of one beta sheet to another is facilitated by seven short loops [2]. Literature reveals that few members of this protein family exhibit association with cancer. In this review, we mainly focus on NGAL (Neutrophil gelatinase-associated lipocalin), an important member of lipocalin protein family and its role in cancer.
NGAL, also known as lipocalin-2 (LCN2), is a 24 kDa secreted glycoprotein in humans encoded by LCN2 gene located on chromosome locus 9q34.11. It was first purified from a culture of mouse kidney cells infected with simian virus 40 (SV40) and subsequently from supernatant of human neutrophils [3]. NGAL is a small molecule which binds and aids in the transport of hydrophobic ligands. It plays a significant role in generating innate immune response and safeguards against bacterial infections by sequestering iron [4]. In addition to ligand binding, it is also involved in proteinprotein interactions and hence, sometimes appears to be complexed with matrix metalloproteinase-9 (MMP9). Depending on the free state or bound state of this glycoprotein, it is termed as “apo” or “holo” NGAL respectively. It is expressed in neutrophils, kidney, prostate, epithelia of the respiratory and alimentary tracts and is majorly used as a biomarker in kidney injury [5,6]. The putative mouse and rat homologues of NGAL are 24p3 and neu-related lipocalin (NRL) respectively. Mouse 24p3 is synthesized in the liver and exerts elevated expression in response to inflammation [7]. A comparison between amino acid sequence of NGAL homologues in different species revealed that human NGAL is highly similar to chimpanzees (98% identity) than mouse and rat (62% and 63% identity respectively) [2].
NGAL is also expressed in several adult normal tissues like breast ducts, liver, lungs, trachea, small intestine, bone marrow, thymus, adipose tissue and macrophages. Negligible expression of NGAL is observed in the normal pancreas, endometrial glands and peripheral blood leukocytes. However, NGAL is completely absent in brain, heart, skeletal muscle, spleen, testes, colon and ovary [2].
NGALR- Receptor NGAL
NGALR is highly conserved in human and is predicted to have twelve transmembrane helices [8,9]. A murine NGAL receptor known as 24p3R was first isolated from FL5.12 cells. This receptor binds to both apo and holo NGAL and enables their internalization. Although, 24p3R internalizes both apo and holo NGAL by endocytosis, interestingly each results in totally different biological processes. The internalization of holo-NGAL by 24p3R receptor donates iron to the cells and thus prevents apoptosis successfully, whereas in the case of apo-NGAL it creates an iron efflux inducing apoptosis [8,9].
Role of NGAL in malignancy
Overexpression of NGAL was first identified in SV40 tumour virus infected quiescent mouse primary kidney cells [10]. It is also referred as protumorigenic protein and, induces tumor progression and chemoresistance in cancer cells. Recently, it has emerged as a biomarker for several benign and malignant diseases. However, it showed differential expression pattern in different cancers (Figure 1).
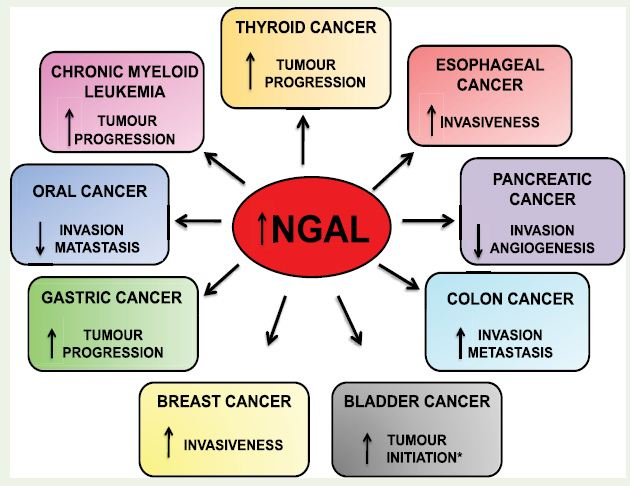
Figure 1: Differential expression and role of NGAL in various cancers; ↑Upregulation,↓Downregulation * in tumor relapse patients reduced levels of NGAL leads to tumor progression
COSMIC database has reported few mutations in LCN2 gene which encodes this protein [13] (Figure 2).
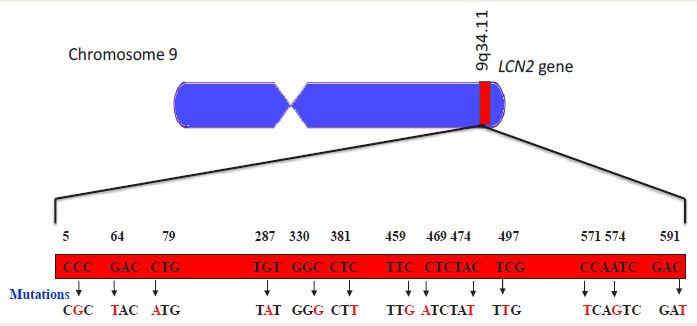
Figure 2: Mutations detected in the gene LCN2. Data based on cosmic database [13].
NGAL in breast cancer
Breast cancer represents the most prevalent type of malignancy in women and led to 14% of the total cancer deaths in 2008 [14]. Despite the significant advances made by constant investigation in the development of biomarkers for the diagnosis of breast cancer, the severity of the disease remains the same. Recently, NGAL was detected in the urine sample of breast cancer patients and has been identified as a biomarker for this cancer [15,16]. In the urine of breast cancer patients, NGAL was found to be complexed with MMP-9, protecting it from degradation and thereby preserving its enzymatic activity [17,18]. This urinary detection of MMP-9/NGAL might open new avenue for non-invasive prediction of disease status in breast cancer patients [18]. MMP9/NGAL complex in the urine of breast cancer patients showed no relationship with the tumor size, TNM stage, patient’s age, menopause status and ER status, but it showed a strong correlation with lymph node metastasis and can beused as a prognostic factor in early screening of disease status [19]. Provatopoulou et al., 2009 [16] reported the presence of NGAL and MMP9 in the serum of breast cancer patients and concluded that it can be used in non-invasive monitoring of breast cancer progression, supporting its role as a biomarker in breast cancer. In a study, MMTV-ErbB2 (V664E) mice lacking LCN2 gene showed notable delay in mammary tumor formation, metastasis and reduced MMP9 activity in blood [20]. Moreover, NGAL expression was enhanced by HER2/phosphoinositide 3-kinase/AKT/NF-κB pathways [20,21]. Reporter assays of full-length or deletion constructs of the NGAL promoter suggested that the production of NGAL is STAT3 and C/ EBPβ dependent [22]. The dependency of NGAL on C/EBPβ was confirmed as the overexpression of C/EBPβ downregulated NGAL and inhibited the migration and invasion of breast cancer [23].These studies indicate that NGAL plays a crucial role as a biomarker in breast cancer and also imply its significance in increasing the invasiveness of this disease.
NGAL in colorectal cancer
Colorectal cancer is the third most common cancer in the world. It is equally prevalent in both men and women and accounts for 9% of total cancer incidences [24]. There are many proteins that are responsible for driving the progression of this disease to the metastatic stage. NGAL is one such protein that is overexpressed and led to the poor prognosis of colorectal cancer patients [25,26]. High expression of NGAL was observed in both colorectal carcinoma cell lines and xenograft mouse models of this disease [26,27]. Plasma NGAL (pNGAL) levels were remarkably higher in colorectal cancer patients when compared to healthy individuals. This elevated pNGAL was associated with higher tumor volume, characteristics of invasion and recurrence, evidencing its prognostic utility in metastatic colorectal cancer patients [28]. In colon carcinoma cell lines, overexpression of NGAL led to subcellular localization of E-cadherin and catenins, decreased E-cadherin mediated cell-cell adhesion, enhanced cell-matrix attachment, increased cell motility and in vitro invasion while downregulation of NGAL entailed reduced in vitro invasion [26]. Contrastingly, KM12SM- a highly metastatic human colon cancer cell line showed decreased expression of NGAL while ectopically expressing, it suppressed in vitro invasion and repressed liver metastasis in BALB/c nude mice [29]. In infectious colitis, downregulation of PPARγ led to the accumulation of lipocalin-2, hence stabilizing MMP9 causing stern colitis. Moreover, mice lacking LCN2 gene stayed protected even at the later stages of infection [30].
However, NGAL cannot be used as a biomarker for determining the disease progression in colorectal cancer as the marked difference in its expression was found only in its early stages [31].
NGAL in esophageal squamous cell carcinoma
Esophageal squamous cell carcinoma is the seventh most common cancer in the world. Upregulation of NGAL led to the activation of differentiation pathway and invasive progression of esophageal squamous cell carcinoma [32]. Zhang et al., 2007 [32] immunohistochemically stained the tissues of different stages of ESCC and reported a weak positive signal in normal esophageal epithelium and altered expression in dysplasia. However, in esophageal squamous cell carcinoma, NGAL was complexed with MMP9 and the enzymic activity of this complex was much elevated in ESCC than in normal mucosa and significantly correlated with tumor invasion [33]. High expression of NGAL has been observed to be responsible for malignant transformation of human immortalized esophageal epithelial cell line SHEE to esophageal carcinoma cell line SHEEC [33-35]. Blocking the expression of NGAL in SHEEC cell lines suppressed invasion in nude mice [34].
NGALR is also overexpressed in esophageal squamous cell carcinoma [35,36]. Cui et al., 2008 [36] reported a high level of NGALR expression on tumor cell membrane and in the cytoplasm. NGALR hypomethylation contributed to its overexpression inesophageal squamous cell carcinoma [36]. This overexpression of both NGAL and NGALR led to the poor survival of esophageal squamous cell carcinoma patients [37].
NGAL in pancreatic cancer
Pancreatic cancer is one of the main leading causes of death and has the lowest survival rate [38]. The low survival rates are due to difficulty in diagnosis at its early stage as the patients show nonspecific symptoms. This necessitates immediate identification of biomarkers for proper diagnosis of this disease at initial stage. Various reports suggested that NGAL can successfully be used as a biomarker in pancreatic cancer [39,40]. Moniaux et al., 2008 [39] reported that NGAL was upregulated 27 fold in pancreatic cancer cells as compared to normal ductal cells. Differential expression of NGAL in different stages of pancreatic cancer was examined by immunostaining. The results showed that NGAL was strongly upregulated in pancreatic cancer and moderately in pancreatitis, but relatively weak expression was detected in healthy pancreas. However, its expression was observed only in well and moderately differentiated pancreatic adenocarcinoma but not in poorly differentiated carcinomas. Serum NGAL level was found to be high in both pancreatic cancer and acute and chronic pancreatitis patients [39]. In an in vitro study, remarkably high level of expression of NGAL was observed in well to moderately differentiated PaCa cells (AsPC-1, BxPC-3, and Capan-2), whereas in the case of moderate to poorly differentiated PaCa cells (PANC-1 and MIAPaCa-2) expression level of NGAL was nearly undetectable. Overexpression of NGAL reduced focal adhesion kinase (FAK) tyrosine-397 phosphorylation without effecting PaCa cells’ survival, viability and their response to chemotherapeutic drugs in vitro. It also suppressed metastasis, reduced tumor volume and angiogenesis in vivo. These results indicated NGAL to be a potential suppressor of invasion and angiogenesis in advanced pancreatic cancer [41]. Later the same group [42] showed that EGF efficiently blocked NGAL expression through activation of the EGFR signaling pathway, which in turn downregulated E-cadherin along with subsequent reduction in NF-κB activation. It has also been reported that NGAL is responsible for gemcitabine resistance in pancreatic ductal adenocarcinoma [43].
NGAL in thyroid carcinoma
Thyroid cancer is a rare type of cancer originating from follicular or parafollicular thyroid cells. It is much more common in women than in men. Barresi et al., 2012 [44] described NGAL to be a potent marker in the identification of malignant follicular cell derived thyroid tumors. NGAL is highly expressed in the human thyroid carcinoma FRO cell line and other poorly differentiated thyroid cancer cell lines [45]. Iannetti et al., 2008 [45] reported that NF-κB helps in thyroid tumor cell survival by controlling iron uptake via NGAL.
NGAL in chronic myeloid leukemia (CML)
Chronic myeloid leukemia (CML), also known as chronic granulocytic leukemia is a cancer of the white blood cells. Fusion protein BCR/ABL formed due to reciprocal translocation of chromosomes 9 and 22 plays a key role in the development of CML[46]. Levels of NGAL expression is correlated with BCR/ABL induced tumorigenesis. Leng et al., 2008 [47] studied the role of 24p3 (Mouse NGAL) and BCR/ABL in leukemia and reported that the mice receiving both BCR/ABL and 24p3 developed leukemia. In addition, primary stable cell line from wild-type mouse marrow cells expressing BCR/ABL caused solid tumors in nude mice whereas a similar BCR/ ABL+ cell line from 24p3 null mice did not develop any tumor. Plasma levels of NGAL in both leukemic mice and CML patients were high [47]. Similarly, blood and serum of different stages of CML patients showed elevated NGAL mRNA and protein expression [48].
It was also evidenced that BCR/ABL oncoprotein upregulated apo-NGAL and at the same time suppressed the receptor NGAL (NGALR), which resulted in cells unresponsiveness to secreted apo- NGAL. Therefore, treating the cells with BCR/ABL inhibitor- imatinib upregulated NGALR resulting in apo-NGAL mediated apoptosis [8].
NGAL in oral cancer
Oral cancer is the sixth most common cancer in the world. The 5 year survival rate of this cancer is only 50%. The death rate due to this cancer has been increasing tremendously over past 30 years. So there is an urgent need for the development of novel biomarkers for early diagnosis of this fatal disease. NGAL holds enormous prospects to serve as a biomarker with an expression profile associated with the degree of differentiation of tumors. Interestingly, it was found that the expression of NGAL decreases from well differentiated to moderatelydifferentiated and completely lost in poorly differentiated tumors [49]. Similar results have also been observed in oral cancer cell lines of different grades of invasiveness. Expression of NGAL was found to be high in weakly invasive (SCCKN, HSC-2 and OSC-20), weak in mildly invasive (HSC-3, OSC-19, and SCC-25) and negative in highly invasive (HOC-313 and TSU) oral squamous cell carcinoma cell lines. Moreover, the expression of NGAL was found to be inversely correlated with E-cadherin and MMP-9 [49].
NGAL in other common cancers
In addition to the above mentioned cancers, expression of NGAL was studied in other commonly occurring cancers like kidney tumors, hepatocellular carcinoma, lung cancer, endometrial cancer and gastric cancer. In hepatocellular carcinoma, overexpression of NGAL and/ or NGALR resulted in poor prognosis and poor survival. The overexpression of NGALR can serve as an independent prognostic factor for this disease [50]. In the case of non-small cell lung cancer, NGAL was responsible for the development of erlotinib resistance [51]. In A549 lung cancer cell line, NGAL served as a survival factor by reducing PDK1 inhibitor induced cell death [52]. The expression of NGAL was also increased with the progression of endometrial cancer [53]. In bladder cancer, high expression of NGAL was detected in the urine of patients. Elevated mRNA transcript and protein levels were found in bladder cancer tissues compared to normal tissues [54,55]. In inorganic arsenic (iAs) treated urothelial cells and bladder cancer tissues expression of LCN2 was enhanced by promoter hypomethylation and mutations at the binding sites for NF-κB and c/eBP-α remarkably decreased LCN2 promoter activity [56]. Moreover, overexpression of LCN2 in inorganic arsenic treated urothelial cells enhanced their oncogenic potential. Thus, NGAL offers beneficial prospect to be used as an early diagnostic marker in bladder cancer. High expression of NGAL was observed in advanced gastric cancer patients resulting in poor prognosis [57]. Du et al., 2011 [58] studied the mechanism of expression of NGAL in gastric cancers before and after 12-O-tetradecanoylphorbol-13-acetate (TPA) induction. Overexpression of NGAL upon induction with TPA evidenced the binding of C/EBPβ to the TPA responsive element (TRE) which is present upstream to the transcription initiation site of NGAL thereby boosting its mRNA level [58].
Conclusion
Despite the phenomenal development made in the field of cancer diagnostics and therapy through continual investigation and appraisal, there is no significant improvement in the survival rate of cancer patients. Lack of biomarkers for the early detection of disease, tumor relapse and chemoresistance are the major factors that contribute to the poor prognosis. It is well established that alterations in the expression of many proteins are responsible for the development of cancer. NGAL is one such vital protein which plays a crucial role in various cellular processes and can be successfully used as a biomarker. Elevated expression of NGAL increases the invasiveness of certain cancers, on the contrary, decreases the invasiveness of some other cancers. It also serves as a biomarker and usually detected in the urine of patients with breast and kidney tumors. However, therole of this protein in some cancers is poorly understood. Therefore, deciphering the role of NGAL can provide new insights to the cancer diagnosis and treatment.
Acknowledgement
This work is supported by SB/FT/LS-411/2012, Science andEngineering Research Board (SERB), Department of Science andTechnology (DST), Govt. of India, awarded to Dr. Ajaikumar B.Kunnumakkara.
References
- Flower DR (1996) The lipocalin protein family: structure and function. Biochem J 318 (Pt 1): 1-14.
- Chakraborty S, Kaur S, Guha S, Batra SK (2012) The multifaceted roles of neutrophil gelatinase associated lipocalin (NGAL) in inflammation and cancer. Biochim Biophys Acta 1826: 129-169.
- Kjeldsen L, Johnsen AH, Sengeløv H, Borregaard N (1993) Isolation and primary structure of NGAL, a novel protein associated with human neutrophil gelatinase. J Biol Chem 268: 10425-10432.
- Yang J, Goetz D, Li JY, Wang W, Mori K, et al. (2002) An iron delivery pathway mediated by a lipocalin. Mol Cell 10: 1045-1056.
- Cowland JB, Borregaard N (1997) Molecular characterization and pattern of tissue expression of the gene for neutrophil gelatinase-associated lipocalin from humans. Genomics 45: 17-23.
- Devarajan P (2008) Neutrophil gelatinase-associated lipocalin (NGAL): a new marker of kidney disease. Scand J Clin Lab Invest Suppl 241: 89-94.
- Friedl A, Stoesz SP, Buckley P, Gould MN (1999) Neutrophil gelatinase-associated lipocalin in normal and neoplastic human tissues. Cell type-specific pattern of expression. Histochem 31: 433-441.
- Devireddy LR, Gazin C, Zhu X, Green MR (2005) A cell-surface receptor for lipocalin 24p3 selectively mediates apoptosis and iron uptake. Cell 123: 1293-1305.
- Richardson DR (2005) 24p3 and its receptor: dawn of a new iron age? Cell 123: 1175-1177.
- Hraba-Renevey S, Turler H, Kress M, Salomon C, Weil R (1989) SV40-induced expression of mousegene 24p3 involves a post-transcriptional mechanism. Oncogene 4: 601-608.
- Moniaux N, Chakraborty S, Yalniz M, Gonzalez J, Shostrom VK, et al. (2008) Early diagnosis of pancreatic cancer: neutrophil gelatinase-associated lipocalin as a marker of pancreatic intraepithelial neoplasia. Br J Cancer 98: 1540-1547.
- Leung L, Radulovich N, Zhu CQ, Organ S, Bandarchi B, et al. (2012) Lipocalin2 promotes invasion, tumorigenicity and gemcitabine resistance in pancreatic ductal adenocarcinoma. PLoS One 7: e46677.
- COSMIC: Catalogue Of Somatic Mutations In Cancer.
- Jemal A, Bray F, Center MM, Ferlay J, Ward E, et al. (2011) Global cancer statistics. CA Cancer J Clin 61: 69-90.
- Wenners AS, Mehta K, Loibl S, Park H, Mueller B, et al. (2012) Neutrophil gelatinase-associated lipocalin (NGAL) predicts response to neoadjuvant chemotherapy and clinical outcome in primary human breast cancer. PLoS One 7: e45826.
- Provatopoulou X, Gounaris A, Kalogera E, Zagouri F, Flessas I, et al. (2009) Circulating levels of matrix metalloproteinase-9 (MMP-9), neutrophil gelatinase-associated lipocalin (NGAL) and their complex MMP-9/NGAL in breast cancer disease. BMC Cancer 9: 390.
- Yan L, Borregaard N, Kjeldsen L, Moses MA (2001) The high molecular weight urinary matrix metalloproteinase (MMP) activity is a complex of gelatinase B/MMP-9 and neutrophil gelatinase-associated lipocalin (NGAL). Modulation of MMP-9 activity by NGAL. J Biol Chem 276: 37258-37265.
- Fernández CA, Yan L, Louis G, Yang J, Kutok JL, et al. (2005) The matrixmetalloproteinase 9/neutrophil gelatinase-associated lipocalin complex plays a role in breast tumor growth and is present in the urine of breast cancer patients. Clin Cancer Res 11: 5390-5395.
- Shen ZZ, Zhao W, Gu J, Zhang ZQ, Yan L (2003) Expression of matrix metalloproteinase-9 and its complex in the urine of breast cancer patients. Zhonghua Wai Ke Za Zhi 41: 817-819.
- Leng X, Ding T, Lin H, Wang Y, Hu L, et al. (2009) Inhibition of lipocalin 2 impairs breast tumorigenesis and metastasis. Cancer Res 69: 8579-8584.
- Li SH, Hawthorne VS, Neal CL, Sanghera S, Xu J, et al. (2009) Upregulation of neutrophil gelatinase-associated lipocalin by ErbB2 through nuclear factor-kappaB activation. Cancer Res 69: 9163-9168.
- Jung M, Weigert A, Tausendschön M, Mora J, Ören B, et al. (2012) Interleukin-10-induced neutrophil gelatinase-associated lipocalin production in macrophages with consequences for tumor growth. Mol Cell Biol 32: 3938-3948..
- Wang L, Li H, Wang J, Gao W, Lin Y, et al. (2011) C/EBP ζ targets to neutrophil gelatinase-associated lipocalin (NGAL) as a repressor for metastasis of MDA-MB-231 cells. Biochim Biophys Acta 1813: 1803-1813.
- Haggar FA, Boushey RP (2009) Colorectal cancer epidemiology: incidence, mortality, survival, and risk factors. Clin Colon Rectal Surg 22: 191-197.
- Barresi V, Reggiani-Bonetti L, Di Gregorio C, Vitarelli E, Ponz De Leon M, et al. (2011) Neutrophil gelatinase-associated lipocalin (NGAL) and matrix metalloproteinase-9 (MMP-9) prognostic value in stage I colorectal carcinoma. Pathol Res Pract 207: 479-486.
- Hu L, Hittelman W, Lu T, Ji P, Arlinghaus R, et al. (2009) NGAL decreases E-cadherin-mediated cell-cell adhesion and increases cell motility and invasion through Rac1 in colon carcinoma cells. Lab Invest 89: 531-548.
- Sun Y, Yokoi K, Li H, Gao J, Hu L, et al. (2011) NGAL expression is elevated in both colorectal adenoma-carcinoma sequence and cancer progression and enhances tumorigenesis in xenograft mouse models. Clin Cancer Res 17: 4331-4340.
- 28.Martà J, Fuster J, Solà AM, Hotter G, Molina R, et al. (2013) Prognostic value of serum neutrophil gelatinase-associated lipocalin in metastatic and nonmetastatic colorectal cancer. World J Surg 37: 1103-1109.
- Lee HJ, Lee EK, Lee KJ, Hong SW, Yoon Y, et al. (2006) Ectopic expression of neutrophil gelatinase-associated lipocalin suppresses the invasion and liver metastasis of colon cancer cells. Int J Cancer 118: 2490-2497.
- Kundu P, Ling TW, Korecka A, Li Y, D'Arienzo R, et al. (2014) Absence of intestinal PPARγ aggravates acute infectious colitis in mice through a lipocalin-2-dependent pathway. PLoS Pathog 10: e1003887.
- McLean MH, Thomson AJ, Murray GI, Fyfe N, Hold GL, et al. (2013) Expression of neutrophil gelatinase-associated lipocalin in colorectal neoplastic progression: a marker of malignant potential? Br J Cancer 108: 2537-2541.
- Zhang H, Xu L, Xiao D, Xie J, Zeng H, et al. (2007) Upregulation of neutrophil gelatinase-associated lipocalin in oesophageal squamous cell carcinoma: significant correlation with cell differentiation and tumour invasion. J Clin Pathol 60: 555-561.
- Xu LY, Li EM, Xiong HQ, Cai WJ, Shen ZY (2001) Study of neutrophil gelatinase-associated lipocalin (NGAL) gene overexpression in the progress of malignant transformation of human immortalized esophageal epithelial cell. Prog Biochem Biophys 28: 839-843.
- Li EM, Xu LY, Cai WJ, Xiong HQ, Shen ZY, et al. (2003) Functions of neutrophil gelatinase-associated lipocalin in the esophageal carcinoma cell line SHEEC. Acta Biochim Biophys Sin 35: 247-254.
- Lin JL, Xu LY, Li EM, Cai WJ, Niu YD, Fang KY, Xiong HQ, Shen ZY, Zeng Y (2004) Antisense blocking of NGAL gene expression affects the microï¬lament cytoskeleton in SHEEC esophageal cancer cells. Prog Biochem Biophys 31: 409-415.
- Cui L, Xu LY, Shen ZY, Tao Q, Gao SY, et al. (2008) NGALR is overexpressed and regulated by hypomethylation in esophageal squamous cell carcinoma. Clin Cancer Res 14: 7674-7681.
- Du ZP, Lv Z, Wu BL, Wu ZY, Shen JH, et al. (2011) Neutrophil gelatinase-associated lipocalin and its receptor: independent prognostic factors of oesophageal squamous cell carcinoma. J Clin Pathol 64: 69-74.
- Beger HG, Rau B, Gansauge F, Leder G, Schwarz M, et al. (2008) Pancreatic Cancer - Low Survival Rates. Bertram Poch Dtsch Arztebl Int 105: 255-262.
- Moniaux N, Chakraborty S, Yalniz M, Gonzalez J, Shostrom VK, et al. (2008) Early diagnosis of pancreatic cancer: neutrophil gelatinase-associated lipocalin as a marker of pancreatic intraepithelial neoplasia. Br J Cancer 98: 1540-1547.
- Kaur S, Chakraborty S, Baine MJ, Mallya K, Smith LM, et al. (2013) Potentials of plasma NGAL and MIC-1 as biomarker(s) in the diagnosis of lethal pancreatic cancer. PLoS One 8: e55171.
- Tong Z, Kunnumakkara AB, Wang H, Matsuo Y, Diagaradjane P, et al. (2008) Neutrophil gelatinase-associated lipocalin: a novel suppressor of invasion and angiogenesis in pancreatic cancer. Cancer Res 68: 6100-6108.
- Tong Z, Chakraborty S, Sung B, Koolwal P, Kaur S, et al. (2011) Epidermal growth factor down-regulates the expression of neutrophil gelatinase-associated lipocalin (NGAL) through E-cadherin in pancreatic cancer cells. Cancer 117: 2408-2418.
- Leung L, Radulovich N, Zhu CQ, Organ S, Bandarchi B, et al. (2012) Lipocalin2 promotes invasion, tumorigenicity and gemcitabine resistance in pancreatic ductal adenocarcinoma. PLoS One 7: e46677.
- Barresi V, Vitarelli E, Reggiani Bonetti L, Tuccari G, Barresi G (2012) Diagnostic value of neutrophil gelatinase-associated lipocalin (NGAL) immunoexpression in follicular-patterned lesions of the thyroid gland. Virchows Arch 460: 319-325.
- Iannetti A, Pacifico F, Acquaviva R, Lavorgna A, Crescenzi E, et al. (2008) The neutrophil gelatinase-associated lipocalin (NGAL), a NF-kappaB-regulated gene, is a survival factor for thyroid neoplastic cells. Proc Natl Acad Sci 105: 14058-14063.
- Melo JV (1996) The molecular biology of chronic myeloid leukaemia. Leukemia 10: 751-756.
- Leng X, Lin H, Ding T, Wang Y, Wu Y, et al. (2008) Lipocalin 2 is required for BCR-ABL-induced tumorigenesis. Oncogene 27: 6110-6119.
- Villalva C, Sorel N, Bonnet ML, Guilhot J, Mayeur-Rousse C, Guilhot F, Chomel JC, Turhan AG (2008) Neutrophil gelatinase-associated lipocalin expression in chronic myeloid leukemia. Leuk Lymphoma. 49: 984-988.
- Hiromoto T, Noguchi K, Yamamura M, Zushi Y, Segawa E, et al. (2011) Up-regulation of neutrophil gelatinase-associated lipocalinin oral squamous cell carcinoma: relation to cell differentiation. Oncol Rep 26: 1415-1421.
- 50.Zhang Y, Fan Y, Mei Z (2012) NGAL and NGALR overexpression in human hepatocellular carcinoma toward a molecular prognostic classification. Cancer Epidemiol 36: e294-299.
- Krysan K, Cui X, Gardner BK, Reckamp KL, Wang X, et al. (2013) Elevated neutrophil gelatinase-associated lipocalin contributes to erlotinib resistance in non-small cell lung cancer. Am J Transl Res 5: 481-496.
- Tong Z, Wu X, Ovcharenko D, Zhu J, Chen CS, et al. (2005) Neutrophil gelatinase-associated lipocalin as a survival factor. Biochem J 391(Pt 2): 441-448.
- Miyamoto T, Asaka R, Suzuki A, Takatsu A, Kashima H, Shiozawa T (2011) Immunohistochemical detection of a specific receptor for lipocalin2 (solute carrier family 22 member 17, SLC22A17) and its prognostic significance in endometrial carcinoma. Exp Mol Pathol. 91: 563-568.
- Candido S, Maestro R, Polesel J, Catania A, Maira F, et al. (2014) Roles of neutrophil gelatinase-associated lipocalin (NGAL) in human cancer. Oncotarget 5: 1576-1594.
- Monier F, Mollier S, Guillot M, Rambeaud JJ, Morel F, et al. (2002) Urinary release of 72 and 92 kDa gelatinases, TIMPs, N-GAL and conventional prognostic factors in urothelial carcinomas. Eur Urol. 42: 356-363.
- Wang HH, Wu MM, Chan MW, Pu YS, Chen CJ, et al. (2014) Long-term low-dose exposure of human urothelial cells to sodium arsenite activates lipocalin-2 via promoter hypomethylation. Arch Toxicol. PMID: 24570342.
- Wang HJ, He XJ, Ma YY, Jiang XT, Xia YJ, et al. (2010) Expressions of neutrophil gelatinase-associated lipocalin in gastric cancer: a potential biomarker for prognosis and an ancillary diagnostic test. Anat Rec (Hoboken) 293: 1855-1863.
- Du ZP, Yuan HM, Wu BL, Chang JX, Lv Z, et al. (2011) Neutrophil gelatinase-associated lipocalin in gastric carcinoma cells and its induction by TPA are controlled by C/EBPβ. Biochem Cell Biol 89: 314-324.