Research Article
Cadmium Toxicity Induces Production of H2O2 and Ethylene in Tomato Suspension Cell Culture
Prasad V1*and Upadhyay RS2
1Institute of Environment and Sustainable Development, Banaras Hindu University, Varanasi, India
2Department of Botany, Institute of Science, Banaras Hindu University, Varanasi, India
Corresponding author: Vishal Prasad, Institute of Environment and Sustainable Development, Banaras Hindu University,Varanasi, India, E-mail: ways2good@gmail.com
Citation:Prasad V, Upadhyay RS. Cadmium Toxicity Induces Production of H2O2 and Ethylene in Tomato Suspension Cell Culture. J Environ Soc Sci.02 2017;4(1): 126.
Copyright © 2017 Prasad V. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Environmental and Social Sciences | Volume: 4, Issue: 1
Submission: 09/03/2017; Accepted: 22/03/2017; Published: 27/03/2017
Abstract
Cadmium (Cd) is a non-essential toxic heavy metal and its interaction with plants causes toxicity to them with an overall reduction in plant productivity. Several physiological and biochemical changes occur when plants are exposed to Cd. The aim of the present work was to investigate the production of H2O2 and ethylene induced by toxicity of Cd in suspension cell culture of tomato. Upon exposure to Cd, a high amount of cell death was observed in suspension cell cultures of tomato. This cell death induced by Cd was accompanied by a significant increase in H2O2 and ethylene production. When inhibitors of H2O2 and ethylene were applied in the suspension cell cultures a significant decrease in the amount of cell death was observed following exposure to Cd. Thus the findings of the present investigation indicate that both H2O2 and ethylene are key players in mounting the toxic effects of Cd in suspension cell cultures of tomato.
Keywords:
Cadmium; Cell death; Inhibitors; Stress; Tomato; Toxicity
Abbreviations
AVG: Aminoethoxy Vinyl Glycine; Cd: Cadmium; Cu: Copper;H2O2: Hydrogen Peroxide; ROS: Reactive Oxygen Species; TCA:Trichloroacetic Acid
Introduction
Plant stress is usually defined as an external factor that exerts a disadvantageous influence on it. Stresses trigger a wide range of plant responses, from altered gene expression and cellular metabolism to changes in growth rates and crop yields. The duration, severity andrate at which a stress is imposed, they all influence a plants responseto a particular stress. A response may be triggered directly by a stressor may result from stress-induced injury. Plants respond to stres through a variety of mechanisms and at times failure to compensate for a severe stress above the threshold or tolerance capacity of the plant may result in plant cell death.
Excess uptake of heavy metal ions into the symplast is deleteriousto most plant species [1,2]. Metal elements play an essential role as micronutrients, for example copper (Cu), or lack an essential function, for example cadmium (Cd).
Cadmium is a non-essential element with negative effects on growth and development of plants. Various sources like nickelcadmium batteries, electroplating process, pigment manufacturing, power stations, metal working industries, urban traffic and cigarette smoke are responsible for its release in the environment [3]. Because of its high toxicity and great solubility in water, Cd is considered asan extremely significant pollutant in the environment [4]. Minerals uptake in plants is hampered by Cd [5]. The presence of Cd also affects many physiological processes like stomatal opening, transpiration and photosynthesis resulting into symptoms like chlorosis, leaf rolls, stunting and even necrosis [3,6,7]. Cadmium inhibits chlorophyll biosynthesis and reduces the activity of enzymes of CO2 fixation process thereby producing alterations in the chlorophyll metabolism [7-9]. Cadmium induces lipid peroxidation and hinders the functionality of membranes [10]. Cadmium toxicity produces oxidative stress by either increasing the production of Reactive Oxygen Species (ROS) or by decreasing the activity of enzymatic and non-enzymatic antioxidants [3,11-13]. The ROS are produced in the cell as by-products of several metabolic steps. Depending on its concentration H2O2 is one such ROS which acts both as toxic by-product of cellular metabolism and as an important signaling molecule [14].
Reactive oxygen species are involved in the response of plants toall kind of biotic as well as a biotic stresses [15,16]. These ROS can trigger unlimited oxidation of various cellular structures, leading tom oxidative destruction of the cell and the common ROS include Singlet Oxygen (1O2), Superoxide Radical (O2•-), Hydrogen Peroxide (H2O2) and Hydroxyl Radical (•OH) [17].
When plants are subjected to Cd stress, a variety of ROS, such assuperoxide, hydrogen peroxide and hydroxyl radicals are generated.These ROS cause oxidative damage in plants [16]. Free radicals are toxic to living organisms unless removed rapidly, destroyed or inactivated by various cellular components. In the absence of effective mechanisms that remove or scavenge free radicals, they can seriously damage plant by lipid peroxidation, protein degradation, breaking of DNA and cell death [18,19].
The simple gaseous olefin ethylene is a classical plant hormone with vital roles in many aspects of plant growth and development [20]. The role of ethylene as a stress hormone during various stressful conditions faced by plants is of prime interest [21].
Ethylene is known to produce serious effects on growth anddevelopment of plants [22,23]. Amongst the best known effect of ethylene is the induction of ripening of fruits, especially those of tomato fruits. Several other processes like seed germination, promotion or inhibition of flowering, abscission of various organs and senescence are also known to be effected by ethylene. Biosynthesis of ethylene is a highly regulated process and is under both positive and negative feedback control [24]. Ethylene production is induced under various external stress conditions such as wounding, pathogen invasion, heavy metal toxicity and flooding [22,25,26]. Ethylene synthesis is also increased upon treatment with Cd, Cu, Fe, Zn, and in the case of Cd and Cu, this increase is due to an upregulation of ACC synthase transcription and enhanced activity [27].
Here in the present study the objective was to evaluate the toxicityof Cd in the suspension cell culture of tomato and its ability to induceH2O2 and ethylene production and the possible involvement of these molecules in the cell death of suspension cell cultures challenged with Cd by application of certain inhibitors.
Materials and Methods
Generation of the suspension cell culture of tomato
Suspension cell culture of tomato (Lycopersicon esculentum cv. CO-3) was generated on MS medium supplemented with 3% sucrose,1 mgL-1 2,4-D and 0.2 mgL-1 BAP from the friable callus obtained from the leaf explants generated on MS medium supplemented with3% sucrose and 2 mgL-1 BAP [28]. The friable callus obtained was suspended in 50 ml of liquid MS basal medium in a 250 ml flask supplemented with appropriate growth regulators and was kept onan orbital shaker at 100 rpm with 14 h light and 10 h dark cycle andtemperature was adjusted to 25±2 °C.
For subculture, 8 days old suspension cell cultures were centrifuged and sedimented down to remove the old medium. The sedimented cells were then resuspended in 15 ml of fresh medium and then 5 ml from this was transferred to a 250 ml flask containing fresh and sterile 45 ml medium. All this process of subculturing was carried out under aseptic conditions under a laminar flow hood. After 5 days of subculture the suspension cell cultures were used for experimental purposes.
Exposure of tomato suspension cell culture to Cadmium
Treatment with the heavy metal Cadmium in form of CdCl2was given to the suspension cell culture of tomato. Stock solution ofCdCl2 was prepared by dissolving appropriate amount of CdCl2 in sterile deionized water to obtain a concentration of 50 mM. From this stock solution different concentrations of CdCl2 were prepared for treatment.
Treatment of the suspension cell culture of tomato was carried out 5 days after subculture. Different concentrations of the Cd (50,100,150,200 and 250 μM) were used for treatment. The required amount of the Cd was added to 50 ml of suspension cell culture under aseptic conditions in a laminar flow hood and the suspension cell culture were then placed back on the shaking incubator for growth. The amount of cell death induced by the Cd treatment in the suspension cell culture was measured 24 h after Cd treatment. Suspension cell cultures kept without any Cd treatment was used as control. In case of treatment of the suspension cell cultures with catalase (50 U/ml; H2O2 inhibitor) and Aminoethoxy Vinyl Glycine (AVG) (10 μM; ethylene biosynthesis inhibitor) these inhibitors were added 10 min before exposure to Cd.
Cell viability staining
For visualizing the effect of treatment with Cd in the suspensioncell culture, the suspension cell cultures were centrifuged at 5000 g for 10 min and then the suspension cell cultures were resuspended in 10ml of 50 mM phosphate buffer (pH 7.5). To 1 ml of this suspension cellculture was added 10 μL of Evans blue stock solution (100 mg Evansblue dissolved in 10 mL sterile distilled water) and was incubated for15 min at room temperature. Following incubation the suspension cell culture was centrifuged at 2000 g. The pellet was washed once with 1 mL of 50 mM phosphate buffer (pH 7.5) to remove any excess Evans blue. The cells were again centrifuged and resuspended in 1 mL of 50 mM phosphate buffer (pH 7.5). Microscopic examination of the cells was carried out under a light microscope. Dead cells stained deep blue where as healthy live cells showed only faint blue coloruation.
Estimation of cell death
To assess the amount of cell death in the suspension cell culturestreated with Cd, the cells were centrifuged at 2000 g for 5 min and the fresh weight was recorded. The cells were then resuspended in 10 mL of 50 mM phosphate buffer (pH 7.5) and 25 μL of Evans blue stock solution was added to it followed by incubation for 15 min at room temperature. After incubation the cells were centrifuged at 2000 g for 5 min and the pelleted cells were washed with sterile distilled water three times for 15 min each to remove any excess and unbound dye. After this the pelleted cells were resuspended in 2 ml of 50% methanol containing 1% (w/v) SDS and the dye bound to dead cells was solubilized at 60 °C for 30 min, this solubilization step was carried twice. Following each solubilization the cells were centrifuged and the resulting supernatants were pooled and were diluted with sterile distilled water to a final volume of 7 mL. The absorbance of this solution was recorded at 600 nm against blank containing distilled water. The control and inhibitor treated suspension cell cultures were also processed in the similar way.
Estimation of H2O2 production
For measurement of the amount of H2O2 in the suspension cell cultures treated with Cd, the suspension cell cultures were harvested by centrifugation at 5000 g for 5 min. Then the amount of H2O2 in the pellet was determined according to Sagisaka [29]. The treated suspension cells were homogenized in 5% cold Trichloroacetic Acid (TCA) and the homogenate was centrifuged at 17000 g for 10 min at 0 °C. The reaction mixture contained 1.6 ml of supernatant, 0.4 ml of 50% TCA, 0.4 ml of 10 mM ferrous ammonium sulphate and 0.2 ml of 2.5 M potassium thiocynate. The absorbance was recorded at 480 nm after 15 min of incubation. The amount of H2O2 was estimated by a calibration curve prepared with known concentrations of H2O2.
Ethylene production assay
Ethylene production was determined as reported by Harrison [30]. In the suspension cell cultures treated with Cd, the amount of ethylene produced was measured by withdrawing 2 ml air samplesfrom the head space of the flask. Air samples were then analyzed for ethylene content on gas chromatograph equipped with a flame ionization detector.
Statistical analysis
All experiments were done thrice in triplicates for each analysis.Data represented here are mean of a single experiment done in triplicates with ±SE represented as bars. Variance analysis (ANOVA) was performed on experimental data and statistical significance at p<0.05 level was judged by the Least Significant Differences (LSD) method. Statistical analysis were performed using statistical program SPSS (SPSS Inc, Chicago, IL, USA).
Results
Cell viability in tomato suspension cell cultures exposed to Cd
Cellular toxicity caused by exposure to Cd was observed by staining with Evans blue. Viable cells were mostly colourless or stained only very faint blue, and exhibited an intact protoplast with dense cytosol (Figure 1A), while a deep blue coloration and protoplast shrinkage were observed in cells challenged with Cd (Figure 1B). The cell death rates were determined using a spectrophotometric assay (Figure 2). A very significant increase in the percentage of dead cells was observed after 24 h of Cd treatment. In the control untreated cells the percentage of dead cells in the population was about 11%. In contrast, the rate of cell death reached 27%, 34%, 42%, 80% and 81% after 24 h of treatment with various concentrations of Cd respectively. Cadmium induced cell death in a concentration dependent manner and based on the results obtained 200 μM of CdCl2 was found and selected for treatment in all further experiments as a very significant amount of cell death (80%) was observed after 24 h of treatment.
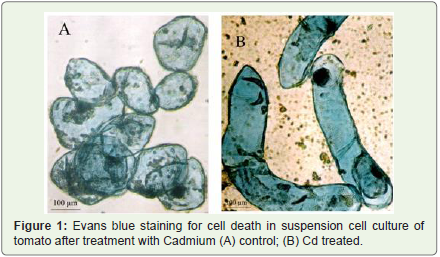
Figure 1: Evans blue staining for cell death in suspension cell culture of tomato after treatment with Cadmium (A) control; (B) Cd treated.
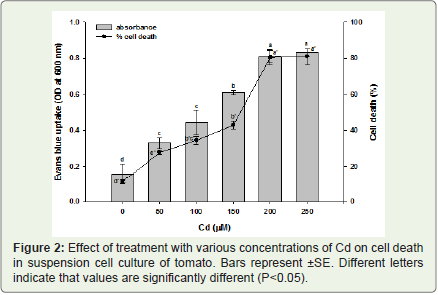
Figure 2: Effect of treatment with various concentrations of Cd on cell death in suspension cell culture of tomato. Bars represent ±SE. Different letters indicate that values are significantly different (P<0.05).
H2O2 production in suspension cell culture following Cd treatment
A gradual increase in H2O2 production was observed in the suspension cell cultures of tomato following exposure to 200 μM Cd (Figure 3). Exposure of the suspension cell culture to Cd stimulated the production of H2O2 by 2 h post treatment. During the peak phase of oxidative burst a very pronounced and large amount of H2O2 was produced between 4-7 h post treatments (Figure 3). In case of controlcells a basal level of H2O2 production was observed which was very insignificant quantitatively in comparison to the Cd exposed ones. Pretreatment of suspension cell cultures with H2O2 scavenger catalase significantly reduced the amount of H2O2 produced in the suspension cell culture and it was very less quantitatively in contrast to the Cd exposed cells (Figure 3).
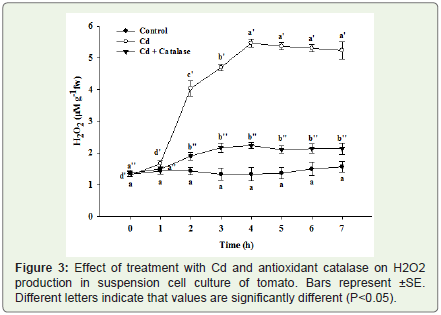
Figure 3: Effect of treatment with Cd and antioxidant catalase on H2O2 production in suspension cell culture of tomato. Bars represent ±SE. Different letters indicate that values are significantly different (P<0.05).
Ethylene production in suspension cell culture following Cd treatment
Ethylene production was measured in the head space volume of the flasks containing the suspension cell cultures of tomato treated with 200 μM Cd, and also in the untreated control suspension cell cultures. Ethylene production was observed to increase sharply on exposure to Cd starting from 2 h post treatment and continued up to 8 h where maximum ethylene production was observed. At 8 h post treatment the amount of ethylene produced was almost 5-6 folds higher than that of control after which ethylene production started declining and a significantly declined production was observed at 14 h post treatment (Figure 4). In case of control cells an extremely lowlevel of ethylene production was observed. Pretreatment of suspension cell culture with ethylene biosynthesis inhibitor AVG significantly reduced the amount of ethylene produced by the suspension cell culture. The amount of ethylene in AVG pretreated suspension cells was quantitatively equivalent to control untreated cells (Figure 4).
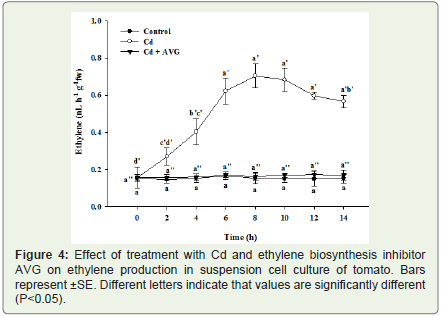
Figure 4: Effect of treatment with Cd and ethylene biosynthesis inhibitor AVG on ethylene production in suspension cell culture of tomato. Bars represent ±SE. Different letters indicate that values are significantly different(P<0.05).
Effect of Inhibitors on Cd toxicity induced cell death
After establishing the production of H2O2 leading to an oxidativestress in the suspension cell cultures of tomato following exposure toCd a potent antioxidant enzyme catalase was applied 10 min before the application of Cd in the suspension cell cultures to asses for a possible role of oxidative burst in the cellular toxicity of Cd inducingcell death. The amount of cell death was reduced to a significant levelfollowing application with 50 U/ml catalase in the suspension cell culture 24 h post treatment and was very much comparable to control cells as was evidenced by lower amount of Evan’s blue uptake (Figure 5).
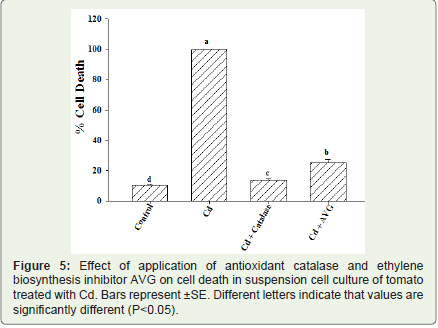
Figure 5: Effect of application of antioxidant catalase and ethylene biosynthesis inhibitor AVG on cell death in suspension cell culture of tomato treated with Cd. Bars represent ±SE. Different letters indicate that values are significantly different (P<0.05).
Similarly it was also observed that exposure of suspension cell cultures to Cd led to ethylene production. To asses for the involvement of ethylene in cellular toxicity of Cd and its possible involvement in the cell death signaling cascade inhibitor of ethylene biosynthesis was tested. Application of 10 μM AVG significantly reduced the amount of cell death induced by Cd exposure 24 h post treatment and the amount of cell death was almost comparable to control where no treatment was given (Figure 5). This inhibition of cell death by application of AVG indicates for possible role of ethylene in the cellular toxicity and cell death cascade induced by Cd in the suspension cell cultures of tomato.
Discussion
The cell morphology of tomato suspension cultures exposed to Cd and stained with Evans blue was screened to assess the toxicity of Cd in the suspension cell population. Dead cells showed typical features of cell death such as cytoplasm shrinkage and nuclear condensation (Figure 1A,Figure 1B). These changes in morphological features of dead cells, severe shrinkage of nuclei and cytoplasm have also been observed earlier in tomato suspension cells treated with CdSO4 and other cell death-inducing chemicals [31,32].
Evans blue uptake method was used to monitor cell death induced by Cd in the treated suspension cell cultures. It was very much apparent from the results obtained that Cd induced a severe cell death in the treated suspension cell cultures as was observed by increased uptake of Evan’s blue (Figure 2). Cadmium induced cell death in a concentration-dependent manner similar to its effect in a number of other plant as well as animal systems.
Independently of the type of stress influence (biotic or abiotic) the accumulation of ROS is an undeniably established fact. It is well known that water deficit, low temperatures, application of pesticides,noxious gasses, harmful radiations, heavy metals, acid rains, high solar irradiation, pathogen attack etc. all cause an increased production oftoxic ROS (O2–, H2O2, OH-, 1O2), which are highly detrimental to all biological systems, i.e. an oxidative stress occurs [33]. Generation of ROS including H2O2 production occurs typically during stress response processes associated with plant-stress interactions.
Treatment with Cd produced an enormous oxidative burst in the suspension cell cultures (Figure 3). Cadmium induced H2O2 production has also been observed in tobacco BY-2 suspension cell cultures [34]. The increased production of ROS is a key event in Cd toxicity in plants and it mediates by inhibition of antioxidant system [35]. Studies have shown that the ROS production induced by Cd is catalyzed by a NADPH oxidase-like enzyme in the plasma membrane [8,34]. The roles of cell wall peroxidases or polyamine oxidases have also been suggested to contribute to H2O2 production [36]. Regardless of the Cd-induced mechanisms of ROS generation, their increased level is always toxic to the system [17,34].
To check whether this oxidative burst was instrumental in cell death cascade induced by the Cd, the antioxidant catalase was applied and then the effect of Cd on H2O2 production and subsequent cell death induction was studied. The application of this antioxidant greatly reduced the amount of H2O2 produced (Figure 3) and also produced a significant decrease in the amount of cell death induced by Cd (Figure 5). This result suggests that oxidative burst is one of the possible causes of cell death induced by Cd in suspension cell culture of tomato. It has been reported earlier that H2O2 scavengers prevented H2O2 accumulation and reduced the symptoms of Cd toxicity in pea leaves [37]. Similarly, cell death inhibition by catalase has been reported in animal cells exposed to Cd. Co-incubation of rat glioma cells with catalase and Cd strongly inhibited Cd-induced DNA ladder formation, indicating that H2O2 is at least partially involved in apoptotic activity of Cd in animal systems [38]. Application of ascorbic acid and catalase has also been found effective in decreasing the amount of cell death induced by Cd in tomato suspension cell cultures [39]. Production of H2O2 was effectively blocked in suspension cells treatment with the antioxidant catalase following exposure to Cd, indicating that the increase in H2O2 is causatively related to Cd toxicity and cell death in tomato suspension cell cultures (Figure 5). Our results are in line with the earlier findings where H2O2 scavengers prevented H2O2 accumulation and reduced the symptoms of Al and Cd toxicity in suspension cell cultures of tomato [32]. Similar effects of Cd on H2O2 production and antioxidant treatment on lowering the H2O2 production was also observed in Arabidopsis leaves [40]. Apart from its role in the induction of cell death H2O2 from the oxidative burst is also a central molecule in coordinating the local and systemic defense signaling. Several unique properties of H2O2, such as its relative stability, ability to diffuse, and ability to cross membranes, enable it to fulfill this role. H2O2 moves from the point of generation forming a concentration gradient, and exhibits different concentration dependent effects. At its threshold concentration H2O2 triggers cell death while lower doses lead to defense activation [14].
Ethylene is a simple gaseous hormone that has been reported toplay an important role in regulating and modulating plant responsesincluding cell death, to both biotic and abiotic stresses [23,41].
Production of the plant phytohormone ethylene was observed to be stimulated by treatment with Cd in the suspension cell culture of tomato (Figure 4). This observation leads to the conclusion that ethylene production is one of the biochemical responses of tomato suspension cell culture following treatment with Cd.
Production of ethylene has also been reported to be enhanced intomato suspension cells treated with CdSO4. To assess the possibility for a role of ethylene in the induction of cell death by Cd, the ethylene biosynthesis inhibitor AVG was applied to check the effect of Cd on cell death induction in suspension cell culture of tomato. Theapplications of AVG lead to a significant reduction in the amountof cell death induced by Cd in the suspension cell culture (Figure 5). This result suggests that ethylene is another major player involved in Cd induced cellular toxicity in suspension cell cultures of tomato.
Cadmium treatment induced a transient increase in ethylene production up to 8 h post treatment (Figure 4). This ethylene apparently plays an important role in Cd toxicity. Suppression ofethylene production with AVG pretreatment reduced the amount of Cd-induced toxicity and cell death in suspension cell culture oftomato (Figure 5). A similar inhibitory effect of AVG was established in Al and Cd induced cell death in tomato suspension cells [32]. Our results are in line with these earlier findings and also show that ethylene production is an important component of heavy metal toxicity in tomato suspension cell cultures.
In a number of other systems like plant-pathogen interactions,following ozone application, treatment of suspension cell culture withfungal toxins and in a diversity of developmental processes, ethylenehas been observed to be closely associated with increased cell death[42,43].
Acknowledgements
The author VP is thankful to Council of Scientific and IndustrialResearch (CSIR), India, for providing financial assistance in form ofSRF during the course of this study.
References
- Clemens S (2001) Molecular mechanisms of plant metal tolerance and homeostasis. Planta 212: 475-486.
- Hall JL (2002) Cellular mechanisms for heavy metal detoxification and tolerance. J Exp Bot 53: 1-11.
- Benavides MP, Gallego SM, Tomaro ML (2005) Cadmium toxicity in plants. Braz J Plant Physiol 17: 21-34.
- Pinto AP, Mota AM, de Varennes A, Pinto FC (2004) Influence of organic matter on the uptake of cadmium, zinc, copper and iron by sorghum plants. Sci Tot Environ 326: 239-247.
- Moreno JL, Hernandez T, Perez A, Garcia C (2002) Toxicity of cadmium to soil microbial activity: effect of sewage sludge addition to soil on the ecological dose. Appl Soil Ecol 21: 149-158.
- Sanita di Toppi L, Gabbrielli R (1999) Response to cadmium in higher plants. Environ Exp Bot 41: 105-130.
- Haouari CC, Nasraoui AH, Bouthour D, Houda MD, Daieb CB, et al. (2012) Response of tomato (Solanum lycopersicon) to cadmium toxicity: Growth, element uptake, chlorophyll content and photosynthesis rate. Afr J Plant Sci 6: 1-7.
- Stobart AK, Griffits WT, Ameen-Bukhari I, Sherwood RP (1985) The effect of Cd2+ on the biosynthesis of chlorophyll in leaves of barley. Physiol Plant 63: 293-298.
- De Filippis LF, Ziegler H (1993) Effect of sublethal concentrations of zinc, cadmium and mercury on the photosynthetic carbon reduction cycle of Euglena. J Plant Physiol 142: 167-172.
- Fodor E, Szabo-Nagy A, Erdei L (1995) The effects of cadmium on the fluidity and H+-ATPase activity of plasma membrane from sunflower and wheat roots. J Plant Physiol 147: 87-92.
- Balestrasse K, Gardey L, Gallego SM, Tomaro ML (2001) Response of antioxidant defence system in soybean nodules and roots subjected to cadmium stress. Funct Plant Biol 28: 497-504.
- Fornazier RF, Ferreira RR, Vitoria AP, Molina SM, Lea PJ, et al. (2002) Effects of cadmium on antioxidant enzyme activities in sugarcane. Biol Plant 45: 91-97.
- Cho U, Seo NH (2004) Oxidative stress in Arabidopsis thaliana exposed to cadmium is due to hydrogen peroxide accumulation. Plant Sci 168: 113-120.
- Gechev TS, Hille J (2005) Hydrogen peroxide as a signal controlling plant programmed cell death. J Cell Biol 168: 17-20.
- Bolwell GP, Bindschedler LV, Blee KA, Butt VS, Davies DR, et al. (2002) The apoplastic oxidative burst in response to biotic stress in plants: a threeâ€component system. J Exp Bot 53: 1367-1376.
- Gallego SM, Benavides MP, Tomaro ML (1996) Effect of heavy metal ion excess on sunflower leaves: evidence for involvement of oxidative stress. Plant Sci 121: 151-159.
- Gzyl J, Rymer K, Gwozdz EA (2009) Differential response of antioxidant enzymes to cadmium stress in tolerant and sensitive cell line of cucumber (Cucumis sativus L.). Acta Biochim Pol 56: 723-727.
- Olmos E, Martinez-Solano JR, Piqueras A, Hellin E (2003) Early steps in the oxidative burst induced by cadmium in cultured tobacco cells (BY-2 line). J Exp Bot 54: 291-301.
- Tian X, Lei Y (2006) Nitric oxide treatment alleviates drought stress in wheat seedlings. Biol Plant 50: 775-778.
- Johnson PR, Ecker JR (1998) The ethylene gas signal transduction pathway: a molecular perspective. Annu Rev Genet 32: 227-254.
- Hall MA, Smith AR (1995) Ethylene and the responses of plants to stress. Bulg J Plant Physiol 21: 71-79.
- Abeles F, Morgan P, Saltveit M (1992) Ethylene in plant biology, 2nd ed. Elsevier Academic Press, 414.
- Upadhyay RS, Prasad V (2008) Programmed cell death in plant-microbe interaction: An overview. J Sci Res 52: 143-149.
- Broekaert WF, Delaure SL, De Bolle MF, Cammue BP (2006) The role of ethylene in host-pathogen interactions. Annu Rev Phytopathol 44: 393-416.
- Arteca RN, Arteca JM (2007) Heavy-metal-induced ethylene production in Arabidopsis thaliana. J Plant Physiol 164: 1480-1488.
- Prasad V, Upadhyay RS (2010) Alternaria alternata f. sp. lycopersici and its toxin trigger production of H2O2 and ethylene in tomato. J Plant Pathol 92: 103-108.
- DalCorso G, Farinati S, Furini A (2010) Regulatory networks of cadmium stress in plants. Plant Signal Behav 5: 663-667.
- Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant 15: 473-497.
- Sagisaka S (1976) The occurrence of peroxide in a perennial plant, populus gelrica. Plant Physiol 57: 308-309.
- Harrison MA (1997) Analysis of ethylene biosynthesis in plant tissue by GC/FID. In: Dashek WV (Ed). Methods in plant biochemistry and molecular biology. CRC Press 153-161.
- Iakimova ET, Woltering EJ, Yordanova ZP (2007) Toxin- and cadmium-induced cell death events in tomato suspension cells resemble features of hypersensitive response. J Fruit Ornam Plant Res 15: 15-19.
- Iakimova ET, Woltering EJ, Kapchina-Toteva VM, Harren FJ, Cristescu SM (2008) Cadmium toxicity in cultured tomato cells -- role of ethylene, proteases and oxidative stress in cell death signaling. Cell Biol Int 32: 1521-1529.
- Noctor G, Foyer CH (1998) Ascorbate and glutathione: keeping active oxygen under control. Annu Rev Plant Physiol Plant Mol Biol 49: 249-279.
- Piqueras A, Olmos E, Martinez-Solano JR, Hellin E (1999) Cd-induced oxidative burst in tobacco BY2 cells: time course, subcellular location and antioxidant response. Free Radic Res 31: S33-S38.
- Schutzendubel A, Schwanz P, Teichmann T, Gross K, Langenfeld-Heyser R, et al. (2001) Cadmium-induced changes in antioxidative systems, hydrogen peroxide content, and differentiation in scots pine roots. Plant Physiol 127: 887-898.
- Apel K, Hirt H (2004) Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annu Rev Plant Biol 55: 373-399.
- Romero-Puertas MC, Rodriguez-Serrano M, Corpas FJ, Gomez M, Del Rio LA, et al. (2004) Cadmium-induced subcellular accumulation of O2–and H2O2 in pea leaves. Plant Cell Environ 27: 1122-1134.
- Watjen W, Beyersmann D (2004) Cadmium-induced apoptosis in C6 glioma cells: influence of oxidative stress. Biometals 17: 65-78.
- Yakimova ET, Kapchina-Toteva VM, Laarhoven LJ, Harren FM, Woltering EJ (2006) Involvement of ethylene and lipid signalling in cadmium-induced programmed cell death in tomato suspension cells. Plant Physiol Biochem 44: 581-589.
- Maksymiec W, Krupa Z (2006) The effects of short-term exposition to Cd, excess Cu ions and jasmonate on oxidative stress appearing in Arabidopsis thaliana. Environ Exp Bot 57: 187-194.
- de J, Yakimova ET, Kapchina VM, Woltering EJ (2002) A critical role of ethylene in hydrogen peroxide release during programmed cell death in tomato suspension cells. Planta 214: 537-545.
- Overmyer K, Brosche M, Kangasjarvi J (2003) Reactive oxygen species and hormonal control of cell death. Trends Plant Sci 8: 335-342.
- Woltering EJ, Yakimova ET, Michaeli R, Kapchina-Toteva VM (2003) Chemical-induced programmed cell death in tomato suspension cells is mediated through ethylene and lipid signalling. In: Ramina A, Chang C, Giovannoni J, Klee H, Perata P, Woltering E (Eds). Advances in plant ethylene research: proceedings of the 7th international symposium on the plant hormone ethylene. Springer pp. 341-351.