Research Article
Screening of Florida Native Green Microalgae as a Potential Source of Biodiesel
Priyanka Narendar1, Krishnaswamy Jayachandran1 and Miroslav Gantar2*
Corresponding author: Miroslav Gantar, Department of Biological Sciences, Florida International University, MMCampus, 11200 SW 8th St. Miami, Fl. 33199,; E-mail: gantarm@fiu.edu
Citation: Narendar P, Jayachandran K, Gantar M. Screening of Florida Native Green Microalgae as a Potential Source of Biodiesel. J Environ Soc Sci.2015;2(1): 111.
Copyright © 2015 Miroslav Gantar et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Environmental and Social Sciences | Volume: 2, Issue: 2
23/04/2015; Accepted: 25/05/2015; Published: 28/05/2015
Abstract
Twenty seven strains of green algae from the Florida aquatic habitats were screened for their ability to accumulate lipids. In addition, Botryococcus braunii was used as the reference strain. This preliminary screening for lipid content was performed with laboratory grown algal cultures using the fluorescent dye Nile red. The lipid accumulation was followed during the course of cultivation as well as under conditions of nitrogen and phosphorus deprivation. Of all strains tested, an increased lipid accumulation was found in five strains (Coelastrum sp. strain 46-4 and strain 108-5, Chlorococcum sp. strain64-12, Dactylococcus sp. strain 64-10 and Stigeoclonium sp. strain 64-8) and they were used for further studies. A linear relationship between algal biomass yield (culture age) and lipid accumulation was observed in most strains tested (Coelastrum sp. strain 46-4 and strain 108-5, Chlorococcum sp. strain 64-12, and Stigeoclonium sp. strain 64-8). Significantly higher lipid accumulation under nitrogen deprivation was observed in only two strains while five strainshad elevated lipid accumulation under phosphorus deprivation. Apparently, nutrient deprivation conditions affect different strains differently. Three strains contained more than 50% of lipid of dry biomass (Coelastrum 108-5, Chlorococcum 64-12 and Stigeoclonium 64-8) and therefore were identified as potentialorganisms to be used in algae-based biodiesel technology.
Keywords: Biofuel; Green algae; Lipids; Nile red
Introduction
The need for renewable energy sources is increasing rapidly as result of rising global population and industrialization. Diminishing of fossil fuel reserves, and increasing concerns about energy security and climate change have led to an increased interest in biofuel esearch [1]. One form of biofuels, which is currently receiving much attention, is biodiesel. Current sources of commercial biodiesel include soybean and palm oil, animal fat and cooking oil waste [2]. The advantage of using biodiesel is that it is a renewable fuel, nontoxic and biodegradable [3]. However, despite the benefits that biodiesel commercialization can provide, the economic aspect of its development at large-scale is limited by the high cost of vegetable oil [4], resulting in a search for alternative sources of feedstock.
Microalgae are a promising alternative sources for biodiesel [5,6].The algae-for-fuel concept started in the 1970s [7] and has recentlybeen re-invigorated with steady increases in energy prices [2]. Using algae for biofuels have several advantages over conventional cropsincluding high growth rates and oil production per area unit [8,9] andthey can be grown in regions of non-arable land, thereby avoidingcompetition with food crops [10]. Algae also use less water thanthe traditional oilseed crops and contribute to sequestration of thegreenhouse gas carbon-dioxide (CO2) [11,12].
Different kind of lipids, hydrocarbons and other oils are producedby different algal species [13]. Some algae can accumulate up to 80%of lipids by weight of dry biomass [14], however the suitability oflipids for fuel conversion depends on species characteristics [1].
In order to develop a technology of producing biodiesel fromalgae, a selection of a productive organism adapted to local conditionsis crucial. Since green algae are ubiquitous and easy to grow theyrepresent the major groups of algae within lipid-producing species[2]. Florida is considered a suitable location for mass cultivation ofalgae and therefore screening of native green algal strains for theirability to accumulate lipids has great potential. In this work the cell lipid accumulation was determined in cultures of different strains ofgreen algae under varying culture conditions.
Materials and Methods
Organisms and culture conditions
Twenty-seven green algal strains (Chlorophyta) that are part ofa culture collection at Florida International University were used inthe study (Table 1). Taxonomic identification of the strains to thegenus level was based on morphological characteristics [15]. As areference strain, we used Botryococcus braunii, which was purchasedfrom University of Texas, Austin. Algal cultures were grown in BG11medium [16] which was buffered with MES buffer(Sigma-Aldrich) atpH 7.20.The cultivation was carried out in laboratory conditions on ashaker with agitation speed of 150 rpm under cool white light (30 μΕm-2 s-1) and temperature of 24°C.
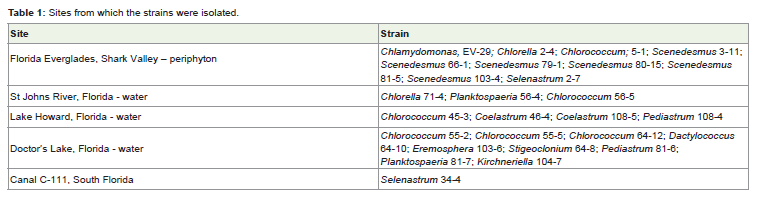
Table 1: Sites from which the strains were isolated.
Growth rate
The growth rate determination was based on dry weight andchlorophyll measurements. For the dry weight measurements, fivemilliliters (three replicas) of samples were filtered onto pre-weighedGF/C (Whatman) filter papers. Filters were dried in an oven at 60°Cto a constant weight and dry weight determined. In the time courseexperiment (Figure 1) the growth rate was determined using changesin chlorophyll content. Five milliliters of culture were centrifuged andthe pellet was extracted with methanol and the absorbance measuredat 666nm and 653nm [17]. The samples were taken every five days.
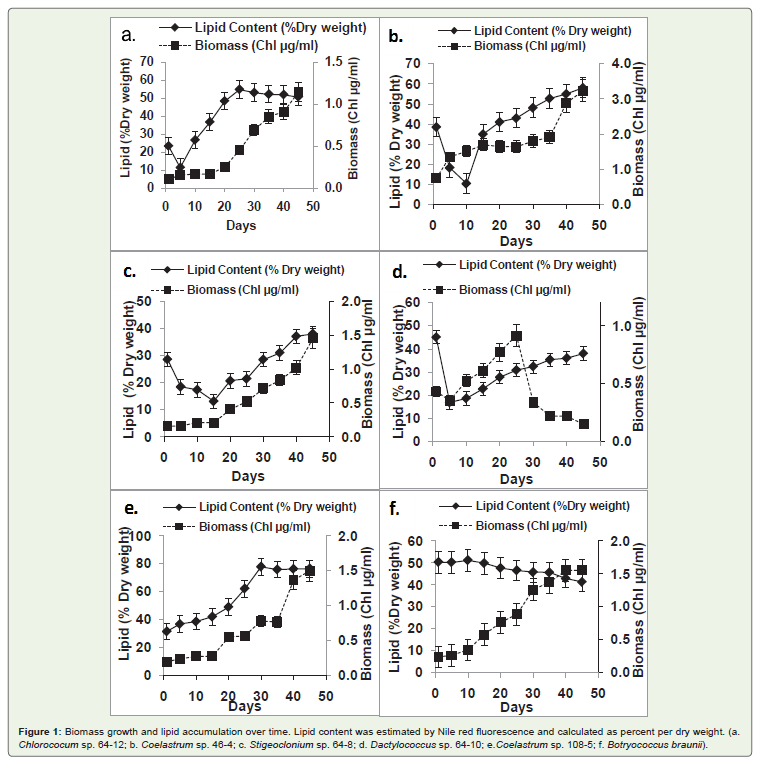
Figure 1: Biomass growth and lipid accumulation over time. Lipid content was estimated by Nile red fluorescence and calculated as percent per dry weight. (a.Chlorococum sp. 64-12; b. Coelastrum sp. 46-4; c. Stigeoclonium sp. 64-8; d. Dactylococcus sp. 64-10; e.Coelastrum sp. 108-5; f. Botryococcus braunii)..
Screening for lipid content
Screening the algal strains for lipid content was based on the Nilered method [18,19]. Algae cultures were grown in 250 ml Erlenmeyerflasks in BG11 medium on a shaker at 25°C under constant light(30μΕm-2s-1). Samples were taken on the 13th and 45th day ofcultivation. Sample of every culture was normalized by dilution toan optical density (OD600) of 0.2. One milliliter of a sample (threereplicates) was transferred into eppendorf tube, homogenized byvortexing, and then DMSO was added (final concentration 20%) toimprove penetration of the dye. To this cell suspension, the Nile redsolution (6.5 mg of Nile red in 26 ml acetone) [20] was added to finalconcentration of 1% (v/v). This mixture was incubated in the dark for10 min and transferred into 96-well plates. The plates were read byusing the excitation wavelength of 530 nm and emission wavelengthat 575 nm [21] on a plate reader Synergy 2 (BioTek, Winooski,VT). The relative fluorescence intensity of Nile red in the stained cells was obtained after subtraction of both the autofluorescence ofalgal cells (cells without Nile red) and the fluorescence of Nile redalone in the medium [20]. These values were normalized by dividingthem by the absorbance values measured at a wavelength of 600nm.In the case of the primary screening (Table 1) the intensity of Nilered fluorescence was further translated into lipid content per 100μl of the culture by using standard curve generated with Triolein asa lipid standard (Fischer Scientific, USA).In other experiments theNile red fluorescence was translated into lipid amount calculated as apercentage of the dry weight.
Triolein standard curve was used in the determination of lipidcontent. One milliliter of triolein which weighed 0.9194g wasdissolved in 9ml of chloroform and diluted to 10-4. The concentrationof the lipid was brought to 90 μg and further diluted in chloroformin order to obtain a concentration in the range of 5 to 50 μg ml-1with the total working standard volume of 3ml. To 3ml of solution,300 ml of chloroform and 150 ml of methanol was added. Also, 7μl of Nile red was added and boiled for 1- 1.5 min and allowed tocool down to room temperature [22]. Fluorescence was read usingthe plate reader at 530nm and 575 nm respectively. The blank in thiscase was chloroform, methanol and Nile red. The calibration curvewas constructed with the fluorescence intensity against the lipidconcentration.
The effect of nitrogen and phosphorus depletion
Those strains showing the highest lipid content were culturedunder varying nitrogen and phosphorous concentrations toprovide information on how different strains respond to nitrogenand phosphorus limitation. Three-week old cultures grown inBG11 medium were centrifuged and the biomass washed with thenitrogen-free or phosphorus-free medium. This biomass was used toinoculate media containing 0%, 50% and 100% of a standard nitrogenand phosphorus content in BG11 medium. The lipid content wasdetermined using the Nile red method for samples taken after 5 and10 days of growth. Nile red fluorescence was translated into percentlipid per unit cell dry weight.
Statistical analysis
All statistical analysis were conducted using SPSS 15.0 software.In this study, it was hypothesized that the lipid content determinedby both gravimetric technique and Nile red method were the same.In order to test our hypothesis, Independent sample T test was usedby comparing the means. It was also hypothesized that nitrogen and phosphorus depletion would increase lipid levels in selected strains.The significance of differences were tested using one way ANOVA.Comparisons that show p-values< 0.05 were considered significant.
Results
Screening the isolates by Nile red method
The initial screening was performed with 27 algal strains. Lipidcontent was estimated during the exponential phase of growth (day13) and at the end of cultivation (day 45). During early exponential phase there was no substantial accumulation of lipidsin any of testedstrains except in reference strain Botryococcus braunii (Table 2). Lipidcontent went up in stationary phase for all the strains. Strains that hadlipid content higher than 27μg per 100μl of culture,were consideredpromising strains and used in further studies includingCoelastrum sp.46-4; Chlorococcum sp. 64-12, Dactylococcus sp. 64-10; Stigeocloniumsp. 64-8; and Coelastrum sp. 108-5, (Table 2). In a separateexperiment using the selected strains, lipid content was determined asa percentage of cell dry weight (Figure 2). Coelastrum sp. strain 46-4had the highest lipid content. After 45 days of cultivation, this straincontained 75% lipids, Coelastrum sp. strain 108-5 and Chlorococcumsp. strain 64-12, contained 73% and 51% of lipids respectively.
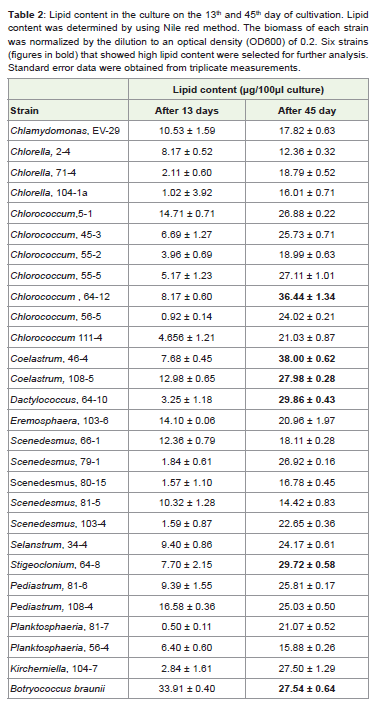
Table 2: Lipid content in the culture on the 13th and 45th day of cultivation. Lipidcontent was determined by using Nile red method. The biomass of each strain was normalized by the dilution to an optical density (OD600) of 0.2. Six strains (figures in bold) that showed high lipid content were selected for further analysis. Standard error data were obtained from triplicate measurements.
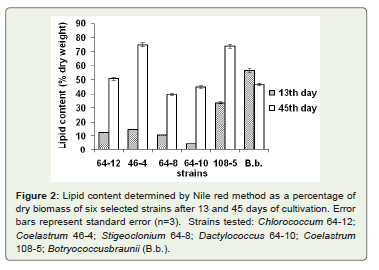
Figure 2: Lipid content determined by Nile red method as a percentage ofdry biomass of six selected strains after 13 and 45 days of cultivation. Errorbars represent standard error (n=3). Strains tested: Chlorococcum 64-12;Coelastrum 46-4; Stigeoclonium 64-8; Dactylococcus 64-10; Coelastrum108-5; Botryococcusbraunii (B.b.).
Lipid accumulation depending on the culture age
In order to determine effect of culture age on lipid accumulation,lipid levelwasdetermined in the biomass of selected six algal strainsevery five days over a period of 45 days. In most tested strains,biomass continuously increased during the entire cultivation period(Figure 2). The exception was Dactylococcus sp. strain 64-10 (Figure 2a) that reached the maximal biomass yield at day 25 followed bya sharp biomass decline. The pattern of lipid accumulation differedfrom one strain to another. In four stains (Chlorococcum sp. strain64-12; Stigeoclonium sp. strain 64-8; Coelastrum sp. strain 46-4; Dactylococcus sp. strain 64-10) there was an initial evident decline inlipid content. After that initial phase, several different patterns of lipid accumulation were observed. There was (1) a continuous increaseof lipid content during the entire cultivation period (Coelastrum sp.strain 46-4; Stigeoclonium sp. strain 64-8; Dactylococcus sp. strain 64-10) (Figure 2b,2c and 2d); (2) the lipid content reached a plateau after25 days of cultivation (Chlorococcum sp. strain 64-12; Coelastrum sp.strain 108-5) (Figure 2a and 2b); (3) and in the case of Botryococcusbraunii, there was no initial change of lipid content with the slowdecline towards the end of cultivation period (Figure f).
A linear regression test showed that only in four strains(Chlorococcum sp. strain 64-12, Coelastrum sp. strain 108-5,Stigeoclonium sp. strain 64-8 and Coelastrum sp. strain46-4) therewas a linear relationship between algal biomass yield and lipidaccumulation with the following correlation coefficients (R2) of 0.834,0.703, 0.632 and 0.612 respectively.
Effect of nitrogen concentration on lipid accumulation
In order to assess effect of nitrogen depletion on lipidlevel, biomass of a 30-day-old culture was washed with nitrogen-free medium andtransferred into nitrogen-free medium and medium containing50% and 100% of nitrogen concentration normally found in BG11medium. The lipid level of the inoculum (day 0) was determined andused as control (Figure 3). Of six strains tested, a significant increase(P > 0.05)in lipid levelin nitrogen-free medium was observed in onlytwo strains. The lipid level increased in Chlorococcum sp. strain 64-12(Figure 3a); and Stigeoclonium sp. strain 64-8 (Figure 3c) 105% and57% respectively. Nitrogen depletion did not significantly affect lipidlevelin other tested strains (Coelastrum sp. strain 108-5; Dactylococcussp. strain 64-10; Coelastrum sp. strain46-4; Botyrococcus braunii).
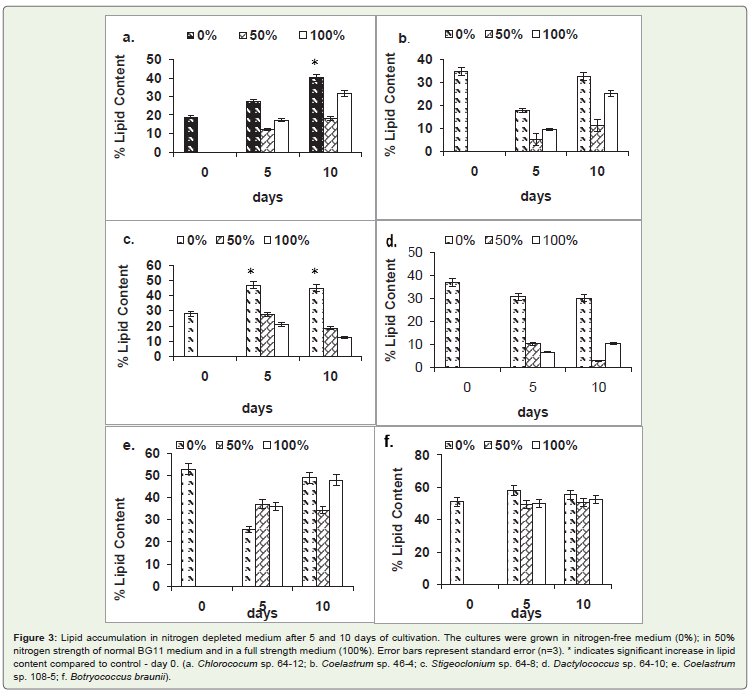
Figure 3: Lipid accumulation in nitrogen depleted medium after 5 and 10 days of cultivation. The cultures were grown in nitrogen-free medium (0%); in 50%nitrogen strength of normal BG11 medium and in a full strength medium (100%). Error bars represent standard error (n=3). * indicates significant increase in lipidcontent compared to control - day 0. (a. Chlorococum sp. 64-12; b. Coelastrum sp. 46-4; c. Stigeoclonium sp. 64-8; d. Dactylococcus sp. 64-10; e. Coelastrumsp. 108-5; f. Botryococcus braunii).
Effect of phosphorous concentrationon lipid accumulation
Of the six strains tested, five showed significant lipid accumulationin phosphorus-free medium after 10 days of cultivation. The highestincrease of lipid level of 116% was observed in Chlorococcum sp. 64-12, followed by Stigeoclonium sp. 64-8(83%); Coelastrum sp. 46-4(58%); Coelastrum sp. 108-5 (30%); and; Botryococcus braunii(26%)(Figure 4c). The only strain that showed decrease of lipid content inphosphorus-depleted medium was Dactylococcus sp. strain 64-10 (Figure 4d). In the strains affected by phosphorus depletion, thesignificant accumulation of lipids occurred after 10 but not after 5days of cultivation. Reduction of phosphorus concentration in themedium by 50% did not affect lipid levels significantly.
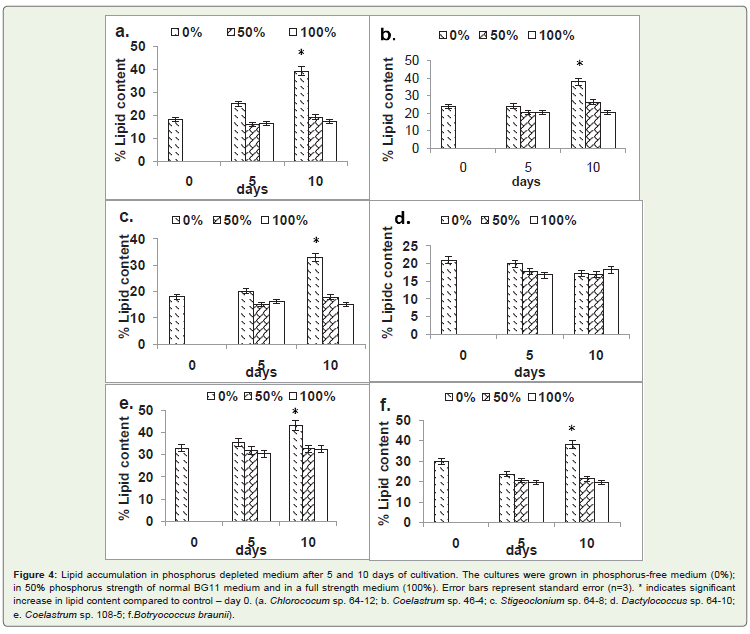
Figure 4: Lipid accumulation in phosphorus depleted medium after 5 and 10 days of cultivation. The cultures were grown in phosphorus-free medium (0%);in 50% phosphorus strength of normal BG11 medium and in a full strength medium (100%). Error bars represent standard error (n=3). * indicates significantincrease in lipid content compared to control – day 0. (a. Chlorococum sp. 64-12; b. Coelastrum sp. 46-4; c. Stigeoclonium sp. 64-8; d. Dactylococcus sp. 64-10;e. Coelastrum sp. 108-5; f.Botryococcus braunii).
Discussion
An earlier study conducted by the Aquatic Species Program [23]emphasized the importance of collecting native algal strains andanalyzing their potential for lipid accretion. Using strains that areadapted to a climate conditions in which they are expected to be masscultured is of crucial significance. In this study, 27 Florida-nativegreen algal (Chlorophyta) strains and their potential to accumulatelipids for the purpose of biodiesel production is investigated. In this initial screening using Nile redmethod, lipid content was determinedin the cultures after 13 and 45 days. The first-time sampling (day 13)was selected based on the study of Kalacheva [24] who reported thatneutral lipids start accumulating after 13 days of cultivation. Fivestrains thatproduced 27 mg of neutral lipids or more per 100 ml ofculture after 45 days of cultivation were used for further analysis(Table 2). In the strains selected for further study, the pattern ofboth growth and lipid accumulation differed from one strain toanother. During 45 days of cultivation, two strains (Coelastrum sp.strain 46-4; Botryococcus braunii) reached stationary phase, threestrains (Chlorococcum sp. strain 64-12; Coelastrum sp. strain 108-5; Stigeoclonium sp. strain 64-8) continued growing till the end ofexperiment and Dactylococcus sp. strain 64-10 showed sharp growthdecline after 30 days.
In most strains tested, the initial lipid content decreased afterinoculation (Figure 2) a consequence of using inoculum in the latestationary phase. This phenomenon of decreasing lipid content upontransferring the cells into new medium has been described beforeand reflects metabolic activity where the lipids are decomposed tosupply energy for cell division and growth [25]. The lipid content inall strains, except control strain Botryococcus braunii, continued toincrease until the end of experiment (45 days). The increase in lipidlevel in aged cultures is in accordance with the report of Chen [18]who showed that the lipid content in the cells of Chlorella vulgarisincreased from 30% to 56% from day 1 to day 12. It is known thatthe old cultures undergo stressful conditions and the majority of thelipids in cells in stationary phase are neutral lipids in the form oftriacylglycerides (TAGs). This is apparently due to the shift in lipidmetabolism from synthesis of membrane to storage lipids eitherthrough de novo biosynthesis or through conversion of membranepolar lipids into TAGs [2].
Botryococcus brauniishowed no increase in lipid level in agingcultures, instead, a decrease of lipid content was observed (Figure 2).Similarly, Alonso [21] reported that culture age had no influence inaccumulation of TAGs in the diatom Phaeodactylum tricornutum.Possible explanation for reduced lipid synthesis in a stationaryphase ofBotyrococcus braunii is that there is an increased rate ofpolysaccharide accumulation [24]. Additionally, Botryococcus brauniihas a slower growth rate than other tested strains. According to Wolf[26], the unusual morphology of this alga could possibly be the factorfor imposing constraints on growth.
The phenomenon of increased lipid level in older cultures is welldocumented [27] and could have important implications on theeconomics of algal biofuels [28]. Selecting strains that have a highgrowth rate and simultaneously capable of accumulating high lipidlevels in a short period of time would be desirable features that couldsignificantly reduce the production costs of biodiesel.
Once a promising strain is selected, ability of the cell toaccumulate lipids can be further improved by manipulating themetabolic pathways through changes in environmental conditions[2]. It is known that under sufficient nutritive conditions, proteinsare synthesized, while during nutrient deprivation, the cell divisionis suppressed and a greater amount of carbon remains available forlipid synthesis [29].
Increased accumulation of lipids, particularly TAGs, has beenobserved in response to nitrogen deficiency in various strains ofmicroalgae [30,31,32]. With nitrogen and phosphorus deprivation,Scenedesmus sp. showed a substantial increase in lipid as high as30% and 53% respectively [33]. This indicates different algal strainsrespond differently to nitrogen deficiency. For example, diatoms donot seem to respond to nitrogen depletion by increasing their lipidcontent [34], while green algae (Chlorophyceae) show differentresponses to nitrogen depletionranging from several fold increases(e.g., Chlorella pyrenoidosa) to no change (Dunaliella species) [35].
Phosphorus deprivation has been reported as another factor thatcontributes to increased lipid accumulation [36,37]. In this study, fiveout of six strains tested responded to phosphorous depletion withan increased accumulation of lipids after 10 days of culturing. Eventhough it is widely accepted that a nitrogen-limitation is the mostimportant factor that induces lipid accumulation [38,39,40], in thiscase, phosphorus deprivation affected most of the strains tested. Inphosphorus-depleted medium, phospholipids of cell membranes arebroken down into neutral lipids [41]. This implies that when usingnutrient deficiency as a method for increasing the lipid synthesisand storage, those conditions should be determined for each strainindividually.
The goal of manipulating algal cell metabolism is to increase lipidcontent without loss of biomass productivity; however, this appearsto be a challenging task. Under nutrient shortage, lipid levels willincrease but cellular growth will decline, resulting in a decreased lipidproduction. Therefore, in order to make microalgae-based biofueltechnology feasible, it is essential to achieve a balance between lipidproduction and biomass yield.
In conclusion, of twenty-seven native Florida algal strainsscreened, three strains contained more than 50% of lipid of drybiomass (Coelastrum 108-5, Chlorococcum 64-12 and Stigeoclonium64-8) and therefore identified as potential organisms to be used inalgae-based biodiesel technology. Deprivation of nutrients such asnitrogen and phosphorus affected various strains differently; moststrains showed better response to phosphorus rather than to nitrogendeprivation. According to this preliminary screening, biodiversityof naturally occurring algae in Florida aquatic environments canprovide a good feedstock in algae-based biodiesel technology.
Acknowledgement
We would like to thank David Berthold for editing our English.
References
- Chisti Y (2007) Biodiesel from microalgae. Biotechnology Advances 25: 294-306.
- Hu Q, Sommerfeld M, Jarvis E, Ghirardi M, Posewitz M, et al. (2008) Microalgal triacylglycerols as feedstocks for biofuel production: perspectives and advances. Plant J 54: 621-639.
- Gerpen JV (2005) Biodiesel processing and production. Fuel Process Technology 86: 1097-1107.
- Antolin G, Tinaut FV, Briceno Y, Castano V, Perez C, et al. (2002) Optimization of biodiesel production by sunflower oil transesterification. Bioresource Technology 83: 111-114.
- Singh A, Nigram PS, Murphy JD (2011) Renewable fuels from algae: An answer to debatable land based fuels. Bioresource Technology 102: 10-16.
- Borowitzka MA, Moheimani NR (2013) Sustainable biofuels from algae. Mitigation and Adaptation Strategies for Global Change 18: 13-25.
- Benemann JR, Goebel RP, Weissman JC, Augenstein DC (1982) Microalgae as a Source of Liquid Fuels. Final Technical Report to US Department of Energy. Washington DC: US Department of Energy.
- Becker EW, Baddiley J, Higgins IJ, Potter WG (1994) Microalgae- Biotechnology and Microbiology. Cambridge University Press, Cambridge.
- Schenk P, Thomas-Hall S, Stephensen E, Marx UC, Mussgnug JH, et al. (2008) Second generation biofuels: High-efficiency microalgae for biodiesel production. BioEnergy Research 1: 20-43.
- Tsukahara K, Sawayama S (2005) Liquid fuel production using microalgae. Journal of The Japan Petroleum Institute 48: 251-259.
- Wang B, Li Y, Wu N, Lan CQ (2008) CO2 bio-mitigation using microalgae. Applied Microbiology and Biotechnology 79: 707-718.
- Papazi A, Makridis P, Divanach P, Kotzabasis K(2008) Bioenergetic changes in the microalgalphotosynthetic apparatus by extremely high CO2 concentrations induce an intense biomassproduction. Physiol Planta132: 338-349.
- Metzger P, Largeau C (2005) Botryococcus braunii: a rich source for hydrocarbons and related ether lipids. Appl Microbiol Biotechnol 66: 486-496.
- Spolaore P, Joannis-Cassan C, Duran E, Isambert A (2006) Commercial applications of microalgae. J Biosci Bioeng 101: 87-96.
- Prescot GW (1962) Algae of the western Great Lake areas. Dubuque, Iowa, W. C. Brown, Co.
- Rippka R, DeReuelles J, Waterbury JB, Herdman M, Stanier RY (1979) Generic assignments, strains histories and properties of pure cultures of cyanobacteria. Journal of General Microbiology 111: 1-61.
- Lichtenthaler HK, Wellburn AR (1983) Determination of Total Carotenoids and Chlorophylls A and B of Leaf in Different Solvents. Biochemical Society Transactions 11: 591-592.
- Chen W, Zhang C, Song L, Sommerfield M, Hu Q (2009) A high throughput Nile red method for quantitative measurement of neutral lipids in microalgae. J Microbiol Methods 77: 41-47.
- Bertozzini E, Galluzzi L, Penna A, Magnani M (2011) Application of the standard addition method for the absolute quantification of neutral lipids in microalgae using Nile red. J Microbiol Methods 7: 17-23.
- Lee SJ, Yoon BD, Oh HM (1998) Rapid method for the determination of lipid from the green alga Botryococcus braunii. Biotechnology Techniques 12: 553-556.
- Alonso DL, Belarbi EH, Fernandez-Sevilla JM, Rodriguez-Ruiz J, Grima EM (2000) Acyl lipid composition variation related to culture age and nitrogen concentration in continuous culture of the microalga Phaeodactylum tricornutum. Phytochemistry 54: 461-471.
- Briaud I, Harmon JS, Kelpe CL, Segu VB, Poitout V (2001) Lipotoxicity of the pancreatic β-Cell is associated with glucose-dependent esterification of fatty acids into neutral lipids. Diabetes 50: 315-321.
- Sheehan J, Dunahay T, Benemann J, Roessler P (1998) A look back at the US Department of Energy’s Aquatic Species Program - Biodiesel from Algae, Report no NREL/TP-580-24190. National Renewable Energy Laboratory, Golden, Colorado.
- Kalacheva GS, ZhilaNO, Volova TG, Gladyshev MI (2002) The Effect of temperature on the lipid composition of the green alga Botryococcus. Mikrobiologiia 71: 286-293.
- Qiao H, Wang G (2009) Effect of carbon source on growth and lipid accumulation in Chlorella sorokiniana GXNN01. Chinese Journal of Oceanography and Limnology 27: 762-768.
- Wolf FR (1983) Botryococcus braunii an unusual hydrocarbon producing alga. Applied Biochemistry and Biotechnology 8: 249-260.
- Greenwell HC, Laurens LML, Shields RJ, Lovitt RW, Flynn KJ (2009) Placing microalgae on the biofuels priority list: a review of the technological challenges. J R Soc Interface 7: 703-726.
- Tran H L, Hong, SJ, Lee CG (2009) Evaluation of extraction methods for recovery of fatty acids from Botryococcus braunii LB 572 and Synechocystis sp. PCC 6803. Biotechnology Bioprocess Engineering 14: 187-192.
- Sukenik A, Wahnon R (1991) Biochemical quality of marine unicellular algae with special emphasis on lipid composition. II. Nannochloropsis sp. Aquaculture 91: 61-72.
- Yeh KL, Chang JS (2011) Nitrogen starvation strategies and photobioreactor design for enhancing lipid production of a newly isolated microalga Chlorella vulgaris esp-31: Implications for biofuels. Biotechnol J 6: 1358-1366.
- Hsieh CH, Wu WT (2009) Cultivation of microalgae for oil production with a cultivation strategy of urea limitation. Bioresource Technology 100: 3921-3926.
- Praveenkumar R, Shameera K, Mahalakshmi G, Akbarsha MA, Thajuddin N (2012) Influence of nutrient deprivations on lipid accumulation in a dominant indigenous microalga Chlorella sp.,bum11008: Evaluation for biodiesel production. Biomass and Bioenergy 37: 60-66.
- Xin L, Hu HY, Ke G, YX Sun (2010) Effects of different nitrogen and phosphorus concentrations on the growth, nutrient uptake, and lipid accumulation of a freshwater microalga Scenedesmus sp. Bioresour Technol 101: 5494-5500.
- Benemann JR, Oswald WJ (1996) Systems and economic analysis of microalgae ponds for conversion of CO2 to biomass. Final report to the Department of Energy. Pittsburgh: Pittsburgh Energy Technology Center, 201 p.
- Borowitzka M (1988) Fats, oils and hydrocarbons. Pp 257-287 in: Borowitzka MA and Borowitzka, LJ eds. Microalgal Biotechnology, Cambridge University Press.
- Roy S (1988) Effects of changes in physiological conditions on HPLC-defined chloropigment composition of Phaeodactylum tncornutum (Bohlin) in batch and turbidostat cultures. Journal of Experimental Marine Biology and Ecology118: 137-149.
- Mandal S, Mallick N (2009) Microalga Scenedesmus obliquus as a potential source for biodiesel production. Appl Microbiol Biotechnol 84: 281-291.
- Evans CT, Ratledge C, Gilbert SC (1985) A rapid screening method for lipid- accumulating yeast using a replica-printing technique. Journal of Microbiological Methods 4: 203-210.
- Roessler PG (1990) Environmental control of glycerolipid metabolism in microalgae: Commercial implications and future research directions. Journal of Phycology 26: 393-399.
- Illman AM, Scragg AH, Shales SW (2000) Increase in Chlorella strains calorific values when grown in low nitrogen medium. Enzyme and Microbial Technology 27: 631-635.
- Beer LL, Boyd ES, Peters JW, Posewitz MC (2009) Engineering algae for biohydrogen and biofuel production. Current Opinion in Biotechnology 20: 264-271.